 2015, Vol. 36
2015, Vol. 36



2. 吉林省农业科学院 农业资源与环境研究中心,吉林 长春 130033
2. Agriculturd Resource and Enviroment Research Institute, Jilin Academy of Agricultural Sciences, Changchun 130033, China
合理增加种植密度是提高大豆产量的重要途径之一,然而随着密度的增加群体内部结构产生了变化,倒伏发生的机率也随之增大.倒伏是大豆高产、稳产和优质的主要限制因子,是控制大豆产量性状的主要数量性状[1].倒伏与品种、气候、土壤条件以及栽培措施密切相关,植株形态特征对抗倒性能有明显影响.作物在弱光等逆境条件下,群体内部出现株高增加、节间伸长、茎粗及干物质积累减少的现象,增加了倒伏风险[2-3].此外,茎秆的化学成分与倒伏的关系密切[4].茎秆强度与木质素和纤维素含量呈正相关[5].倒伏与地上部性状如茎秆强度、茎粗和地下部性状根质量、根长相关[6-7],其中对抗倒伏贡献最大的是茎秆强度,其次是根质量[8].群体内部茎秆和根系的协调生长是减轻倒伏的首要因素[9].磷钾营养与大豆抗倒性及其倒伏程度与产量性状的关系已有一些研究[10-11];钟开珍等[12]和范冬梅等[13]还对大豆种质倒伏性遗传及相关形态的QTL展开了深入研究.而有关大豆不同群体结构下植株综合抗倒性能的研究鲜见报道.因此,通过开展不同种植密度下植株抗倒特性的研究,阐明群体结构与倒伏发生的规律,为实现高种植密度抗倒伏栽培提供理论依据.
1 材料与方法 1.1 材料大豆品种:吉农30,中熟亚有限结荚习性,圆叶、白花,株高90~95 cm,总节数21节;吉农40,中熟亚有限结荚习性,圆叶、紫花,株高90~93 cm,节间短,总节数20节.均由吉林省大豆区域技术创新中心提供.
1.2 试验设计试验于2012年在吉林农业大学大豆区域技术创新中心试验基地进行,土壤为典型黑土,上等肥力水平,碱解氮、速效磷、速效钾分别为120.0、16.5、122.0 mg·kg-1,有机质、全氮、全磷分别为26.90、1.65、0.85 g·kg-1,pH 6.8.采用裂区设计,3次重复,以品种为主区、种植密度为副区,设16、22、28、34、38万株·hm-2共5个密度水平,分别记为M1、M2、M3、M4、M5(理论株距分别为9.6、7.0、5.5、4.5、4.0 cm).每小区5行,行长4.50 m、行距0.65 m,小区面积14.625 m2.施用磷酸二铵和氯化钾分别为150和50 kg·hm-2,播种时全部作为种肥一次施入.于4月29日播种,播种时采用机械开沟、人工条播的方式,将每行所对应的种子量均匀播种在4.50 m行长范围内,各小区出苗后按预设密度定苗,生育中期按常规进行田间管理,2012年9月26日收获测产.M1~M5用种量依文献[14]的公式计算,分别为14.0、19.1、24.4、29.6、33.3粒·m-1.
1.3 测定项目与方法 1.3.1 植株形态指标测定于结荚始期(R3)调查植株形态指标.株高、重心高度、茎粗测定部位为“子叶节”上部1 cm处,基部1~5节节间长和粗(茎秆基部1~5节是影响倒伏的关键节位,其中节间粗测定各节节间中部位置)用卷尺和游标卡尺测定;用天平称节间鲜质量(基部1~5节);计算节间长/粗和单位茎长鲜质量(节间鲜质量/节间长度).
植株重心高度:将植株地上部于子叶节处剪断,之后将保持完整的地上部植株(茎秆的中上部位置)平放于右手食指上,用食指支撑整个植株质量,再于水平方向左右调整植株,当调整到植株与地面处于平行状态时,此时在茎秆上记下食指中心所处位点,使用卷尺测量子叶节至这一位点的距离,即为植株重心高度.
1.3.2 根系伤流量收集及根系指标调查在结荚盛期(R4)将主茎从子叶节处剪断,套上内充脱脂棉的伤流袋,于18:00到次日06:00收集12 h伤流液称质量.之后将地下部根系按照长20 cm、宽20 cm、深40 cm的土体体积挖出,冲洗干净进行根系相关指标的调查.数根瘤数和侧根数(根瘤直径大于0.1 cm、根长大于2 cm);主根长(自子叶节处至主根末端)、主根粗(子叶节下部2 cm处)用卷尺和游标卡尺测定;用天平称根干质量;采用排水法测量根体积.
1.3.3 茎秆压碎强度使用艾力仪器有限公司生产的YYD-1型数字式测力仪测定茎秆压碎强度,将基部1~5各节间的两端放于支撑架凹槽内,然后缓慢向下压,直到茎秆折断为止,此时读出的数值即为该节的压碎强度.计算茎秆抗倒指数,茎秆抗倒指数=压碎强度/重心高度.
1.3.4 倒伏级数调查参照周蓉等[4]的方法调查并加以改进.于开花期、鼓粒期、成熟期调查倒伏发生级数、类型及发生倒伏的部位.0级为小区全部植株直立;1级为轻度倒伏(植株倾斜角≤15°);2级为中度倒伏(15° < 植株倾斜角≤30°);3级为重度倒伏(30° < 植株倾斜角≤45°);4级为严重倒伏(45° < 植株倾斜角≤75°);5级为植株全部匍匐地面(植株倾斜角 > 75°).
1.4 数据分析所有数据使用Microsoft Excel 2003进行处理、SPSS 13.0进行统计与分析(所选数据取各处理平均值,倒伏级数选用R6期数据).
2 结果与分析 2.1 群体密度对植株特征和倒伏发生情况的影响由表 1可知,随着群体密度的增加,株高、重心高度、株高/茎粗逐渐增大;茎粗、茎秆压碎强度、植株抗倒指数逐渐降低.群体的倒伏级数随着密度的增加而升高,说明高密度群体下植株发生倒伏的机率较大.对不同密度处理上述调查指标进行差异显著性分析可知,高密度群体下的各项指标与低密度相比均不同程度达到了显著或极显著水平.在开花期,除M4、M5处理发生0.5级倒伏外,其他处理均未发生倒伏.至鼓粒期,各处理倒伏程度加重,M1和M2处理时倒伏发生级数较低;当种植密度达到28万株·hm-2(M3)时,群体倒伏发生级数明显增加,此时吉农30和吉农40倒伏级数分别为3.33和3.17;由M3到M5时倒伏再次加剧,2个品种倒伏级数最大值均出现在最高种植密度(M5)条件下,田间调查倒伏类型多为茎倒伏.对鼓粒期不同密度的倒伏级数进行差异显著性分析得出,M1、M2分别与M3、M4、M5达到了极显著水平.由表中成熟期各密度的倒伏级数可知,鼓粒期发生轻、中度倒伏的群体在进入成熟期倒伏程度有所减轻,而严重倒伏的变化不大,说明大豆群体在轻、中度倒伏的情况下,植株具备一定的自我恢复能力.
|
|
表 1 不同群体密度下大豆植株特征和倒伏发生情况1) Table 1 The soybean plant characteristics and actual lodging under different planting densities |

随着群体密度的增加,2个品种单株茎秆各节间长度均呈增加趋势,其中第1节长度分别增加1.04和0.90 cm、第3节增加1.39和1.28 cm、第5节增加1.50和2.27 cm.同一种植密度不同节位节间长度随着节位的上升而逐渐增加,其中吉农30在M1、M2处理下第5节比第1节平均长1.86和1.58 cm,而在M4、M5处理下第5节比第1节平均长2.52和2.54 cm,可以看出随着密度的增加其差值逐渐增大(图 1).从图 1中还可看出,高密度条件下的节间长度增长趋势大于低密度的,这一变化趋势在吉农40中表现较为明显,这也是植株高度随密度增加的主要原因.

|
图 1 不同种植密度下大豆茎秆各节位节间长度的变化 Figure 1 The change of internode length under different planting densities M1~M5分别为16、22、28、34、38万株·hm-2. |
由表 2可知,随着群体密度的增加吉农30和吉农40各节位节间粗均呈下降趋势,其中第1节茎粗减少0.30和0.35 cm,第2节茎粗减少0.28和0.35 cm,第3节茎粗减少0.28和0.36 cm,第4节茎粗减少0.26和0.27 cm,第5节茎粗减少0.25和0.24 cm;由此可知,第1~5节位随着密度的增加低节位茎粗较高节位茎粗降低幅度大.由同一节位不同密度下节间粗的变异系数可知,低节位的变异系数大于高节位的变异系数,这也说明群体密度的变化对茎秆低节位的影响较高节位明显.从植株抗倒能力来看,低节位茎粗随密度增加降低较大是植株抗倒能力减弱的原因之一.从第1~5节位茎粗平均值可看出,随着节位的上升节间粗呈先增加后降低的趋势,茎粗最大值出现在第3节.相同种植密度下不同节位茎粗的变异系数表现为高密度大于低密度,说明在高密度群体中茎秆各节节间粗变异较大.
|
|
表 2 不同群体密度下大豆茎秆各节位节间茎粗的变化1) Table 2 The change of internode diameter under different planting densities |

图 2代表不同群体密度下大豆茎秆各节位节间长与节间粗的比值.2个品种第1~5节各节位节间长与节间粗比值随密度增加而逐渐升高,均表现M5 > M4 > M3 > M2 > M1.其中吉农30和吉农40在M2处理下各节位的比值较M1高16.25%和21.28%,M3比值较M1高41.66%和56.17%、M4比值较M1高80.79%和98.57%,M5比值较M1高111.57%和143.75%.相同密度下不同节位节间长与节间粗的比值随着节位的上升节间长与节间粗比值逐渐增加,2个品种均表现第5节 > 第4节 > 第3节 > 第2节 > 第1节.2个品种第2节节间长与节间粗平均比值较第1节高17.11%和-0.18%、第3节较第1节高20.71%和14.57%、第4节较第1节高48.21%和45.17%、第5节较第1节高76.40%和70.18%.随密度的增加吉农40节间长与节间粗比值的增长率较吉农30高,而随着节位的升高吉农40节间长与节间粗比值的增长率较吉农30低.

|
图 2 不同种植密度下大豆茎秆各节位节间长/节间粗的变化 Figure 2 The change of internode length/diameter under different planting densities M1~M5分别为16、22、28、34、38万株·hm-2. |
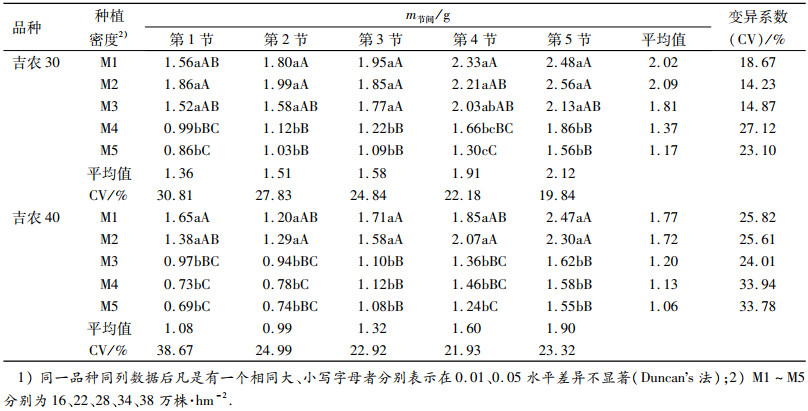
随着群体密度的增加节间鲜质量总体呈降低趋势(表 3),不同密度下吉农40节间鲜质量平均值表现为M1 > M2 > M3 > M4 > M5,而吉农30略有不同,表现为M2 > M1.由各密度下第1~5节节间鲜质量的变异系数(CV)可知,高密度处理的CV大于低密度处理,说明同一植株在高密度条件下各节之间的鲜质量变异程度较低密度大,这可能是影响植株抗倒能力发生变化的原因之一.相同密度下不同节位的节间鲜质量随着节位的上升,节间鲜质量逐渐降低.从各节位对应的CV可知,随着群体密度的增加低节位的CV大于高节位,说明群体密度的改变对茎秆低节位的节间鲜质量影响较大.
|
|
表 3 不同群体密度下大豆茎秆各节位节间鲜质量的比较1) Table 3 Comparation of the fresh mass of soybean internode under different population densities |
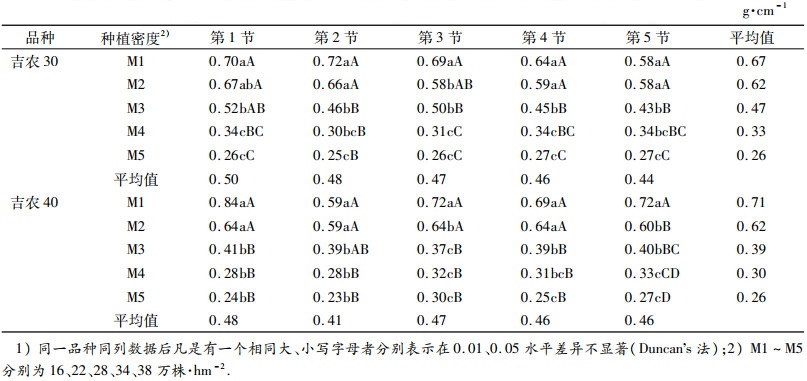
由表 4可知,2个品种各节位单位茎长鲜质量随着群体密度的增加而降低,其中M2各节平均单位茎长鲜质量比M1低7.89%和12.13%、M3比M1低29.30%和45.25%、M4比M1低51.33%和57.15%、M5比M1低60.75%和63.65%.随着群体密度增加, 吉农40单位茎长鲜质量比吉农30降幅大.在低密度处理下,相同密度不同节位的单位茎长鲜质量低节位单位茎长鲜质量大于高节位,高密度下则表现相反.不同群体密度下,低、高节位的单位茎长鲜质量所表现出的逆向变化趋势,将会对植株抗倒能力产生极大的影响.
|
|
表 4 不同密度下茎秆各节位单位茎长鲜质量的比较1) Table 4 The changes of ratio of fresh mass to length of internode under different planting densities |
节间压碎强度是开展作物茎秆抗倒力学性状研究的首要指标.由图 3可知,吉农30和吉农40各节位的节间压碎强度随着群体密度的增加而逐渐降低,其中M2各节位的节间压碎强度的平均值比M1低18.67%和15.26%、M3比M1低30.04%和34.06%、M4比M1低50.01%和57.05%、M5比M1低67.19%和66.75%,随着密度的增加其降幅比值逐渐增大.相同密度下随着茎秆节位的上升节间压碎强度逐渐降低,即相同密度下基本上呈现第1节 > 第2节 > 第3节 > 第4节 > 第5节,2个品种第5节位压碎强度比第1节位分别低39.05%和43.79%.对2品种比较得出,吉农40茎秆压碎强度较吉农30高.

|
图 3 不同种植密度下大豆茎秆各节位节间压碎强度的变化 Figure 3 The change of stalk crushing strength under different planting densities M1~M5分别为16、22、28、34、38万株·hm-2. |
大豆根系所行使的基本功能是获取土壤中作物生长必需的矿物质和水分及固定植株,根系的发育程度直接影响到植株的稳固能力.表 5中单株根瘤数、根干质量、主根粗、主根长、根体积、侧根数和伤流量等性状随群体密度增加而逐渐降低.对吉农30和吉农40的单株根瘤数比较可知,后者的根瘤数大于前者,分析原因可知,大豆的侧生根是根瘤菌着生的主要部位,吉农40的侧根数较吉农30多,侧生根数量多可着生更多的根瘤.当密度由M1增加到M2,单株根干质量、伤流量差异达显著水平,而其他性状差异不显著;密度继续增加到M3时,根系各性状与M1相比均表现出显著或极显著差异.2个品种的根瘤数、主根长、根体积和侧根数在M5与M4间均未达到明显差异.总的来看,除个别处理和性状外,高密度群体下的根系相关性状与低密度相比均达到了极显著水平.
|
|
表 5 不同群体密度下大豆根系相关性状的比较1) Table 5 Comparation of soybean root traits under different planting densities |

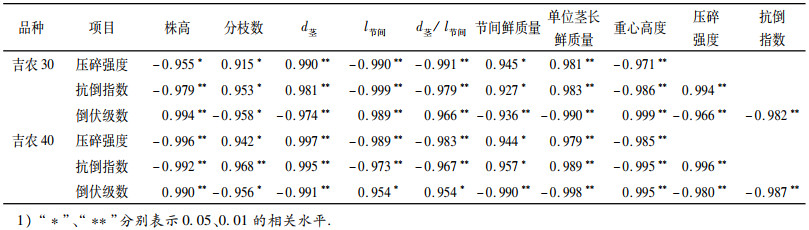
由表 6可知,茎秆压碎强度和抗倒指数呈极显著正相关,而二者与倒伏级数呈极显著负相关.吉农30和吉农40茎秆压碎强度、抗倒指数与其他性状间同样达到显著和极显著水平,因此,可通过这2项指标来衡量植株的抗倒能力.通过相关分析得出,2个品种单株分枝数、茎粗、节间质量和单位茎长鲜质量与倒伏级数呈显著负相关,而与压碎强度和抗倒指数呈极显著正相关;倒伏级数与株高、节间长、节间长/节间粗、重心高度呈极显著正相关,而与压碎强度和抗倒指数呈极显著负相关.
|
|
表 6 倒伏与茎秆性状的相关分析1) Table 6 Correlation analysis of the lodging resistance and stalk characters |
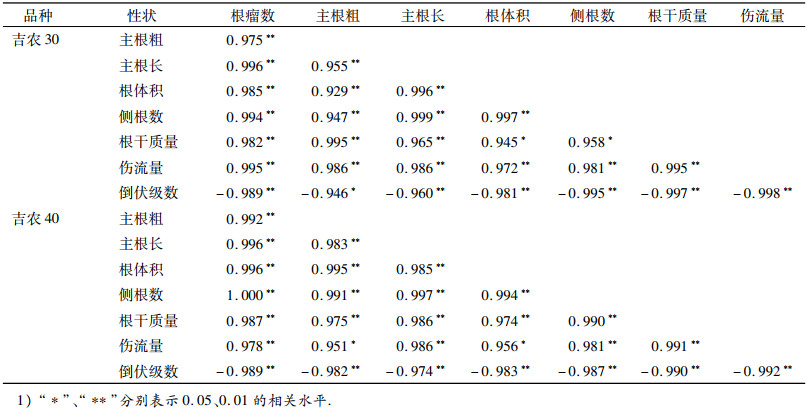
由表 7可知,吉农30和吉农40的倒伏级数与试验中所调查的根系性状均呈负相关,其中与伤流量、根干质量、根瘤数、侧根数等达极显著水平.对各性状相关系数绝对值比较得出,吉农30的倒伏级数与各性状的相关系数依次为:根系伤流量 > 根干质量 > 侧根数 > 根瘤数 > 其他性状;而吉农40有所不同,表现为根瘤数 > 侧根数.所有调查指标中伤流量和根干质量与倒伏级数相关性最大,且与根系其他性状的相关性呈显著或极显著正相关.
|
|
表 7 倒伏与根系性状的相关分析1) Table 7 Correlation analysis between the lodging resistance and root-related traits |
群体中个体发育失调是倒伏发生的内因,抗倒能力与群体的个体质量紧密相关[15].随着种植密度的增加,群体内部对光能资源的竞争逐渐加剧,光照不足成为个体生长发育的首要制约因素.光照作为影响倒伏的重要环境因子之一[16],如不足将会导致茎秆节间伸长,机械组织脆弱、茎秆强度降低、田间倒伏发生严重[2, 6].杨世民等[17]在水稻上研究得出,随密度的增加,茎秆基部节间变细长、茎壁变薄、充实度变差,从而影响茎秆的倒伏指数和抗倒伏能力.本研究得出,大豆株高、重心高度随群体密度增加而升高,茎秆压碎强度、植株抗倒指数逐渐降低,谢甫绨等[18]也得出了同样的结论.倒伏级数随群体密度的增加而增加,植株发生倒伏的风险较大.研究发现,在生育后期,大豆群体发生轻、中度茎倒伏的情况下,植株在临近成熟期时具有一定的自我恢复能力.
植株地上部茎秆和地下部根系性状是影响倒伏的两大因素[13],植株的抗倒伏能力是基部茎秆和根系各性状相互作用的结果,任何一个性状的削弱,都会影响抗倒性.由于植株形态发生变化,如近基部节间伸长,重心高度增加,茎秆粗度、穿刺强度和硬皮组织厚度的降低,致使茎秆抗倒能力差[19, 7].然而,随着株高和重心高度的降低,单茎干物质积累量增多,节间缩短增粗,机械强度提高,田间不易发生倒伏[20].陈小林等[21]指出,随着最长节间下移和节间长度大幅度的增长,植株易倒伏.本研究得出,高密度下的节间长度增长趋势较低密度明显,节间长度的增加是植株增高及重心上移的直接原因;同时得出,随密度增加,低节位茎粗、节间鲜质量降低较高节位明显,茎秆的形态及物质分配发生了改变,导致了茎秆抗倒能力的减弱.刘唐兴等[22]对油菜研究得出,主根长、根粗与侧根数、根体积存在显著正相关;倒伏与根粗存在较强的负相关性,但未达显著水平.沈学善等[23]研究得出,玉米根倒率与单株鲜质量、根系干质量呈显著负相关.本文研究得出,随着群体密度增加,大豆单株伤流量、根干质量、根瘤数、侧根数等显著降低,倒伏逐渐加重.其中,伤流量和根干质量与倒伏级数相关性最大,并且与根系其他性状呈显著或极显著正相关,可选用根系伤流量和干质量作为植株抗倒能力的判定指标.
倒伏与茎秆性状存在明显的相关性.大量研究得出,基部茎秆的倒伏指数与株高、重心高度和基部各节间长度呈正相关[12],而与茎粗、茎质量呈负相关[17],且相关系数大多达到显著或极显著水平[24].茎秆机械强度与基部节间质量呈极显著正相关,与倒伏级数呈极显著负相关[25].本试验中茎秆压碎强度和抗倒指数呈极显著正相关,均与倒伏级数呈极显著负相关.茎秆的压碎强度、抗倒指数与茎秆其他性状间相关达到显著和极显著水平,因此,通过这2项指标来衡量植株的抗倒能力是可行的.
密度改变了植株地上部群体的内部结构,同时也对地下部根系产生了极大的影响,结果导致大豆单株抗倒能力发生变化.地下部根系的根瘤数、根干质量、伤流量和侧根数等随密度增加而降低,倒伏级数与各性状间均呈负相关.随密度增加,植株茎秆压碎强度、抗倒指数显著降低,与倒伏呈极显著负相关;而株高、重心高度、节间长度增加,与倒伏呈极显著正相关.高密度群体结构下,节间长度增长趋势较低密度明显、低节位单位茎长鲜质量小于高节位,而低密度下则表现相反;此外,随密度增加,低节位茎粗、节间鲜质量降低幅度较高节位明显;茎秆第1~5节的物质分配的变化影响了植株的抗倒能力.本研究得出大豆植株抗倒伏的综合性状为:地上部株高适宜、重心较低、近基部节间短粗、基部节间单位茎长鲜质量较高,力学上表现出较强的茎秆抗折力.地下部根系粗壮发达,主根入土较深,根活力旺盛具有较高的伤流量和根干质量,主根着生的侧生根量多、根瘤数量多.本研究是在常规大豆群体中进行的,其部分研究结论同样适用于杂交大豆群体,可为杂交大豆抗倒伏高产栽培提供理论参考.
| [1] |
张丽伟, 齐照明, 刘春燕. 基于Meta分析的大豆倒伏性相关QTL的整合[J]. 植物遗传资源学报, 2010, 11(6): 755-759. (  0) 0) |
| [2] |
CAMPBELL C A, READ D W L. Influence of air temperature, ligintensity and soil moisture on the growth, yield and some grow analysis characteristics of Chinook wheat grown in the grow chamber[J]. Can J Plant Sci, 1968, 48: 299-311. DOI:10.4141/cjps68-053 ( 0) |
| [3] |
陈怀珠, 孙祖东, 杨守臻, 等. 荫蔽对大豆主要性状的影响及大豆耐荫性鉴定方法研究初报[J]. 中国油料作物学报, 2003, 25(4): 78-82. DOI:10.3321/j.issn:1007-9084.2003.04.018 ( 0) |
| [4] |
周蓉, 涂赣英, 沙爱华, 等. 大豆种质的倒伏性调查及其相关农艺性状分析[J]. 大豆科学, 2007, 26(1): 41-44. DOI:10.3969/j.issn.1000-9841.2007.01.010 ( 0) |
| [5] |
郭玉华, 朱四光, 张龙步, 等. 不同栽培条件对水稻茎秆材料学特性的影响[J]. 沈阳农业大学学报, 2003, 34(l): 4-7. ( 0) |
| [6] |
李金才, 尹钧, 魏凤珍, 等. 播种密度对冬小麦茎秆形态特征和抗倒指数的影响[J]. 作物学报, 2005, 31(5): 662-666. DOI:10.3321/j.issn:0496-3490.2005.05.023 ( 0) |
| [7] |
崔海岩, 靳立斌, 李波, 等. 遮阴对夏玉米茎秆形态结构和倒伏的影响[J]. 中国农业科学, 2012, 45(17): 3497-3505. ( 0) |
| [8] |
韩清瑞. 冬小麦基部节间的形态结构与倒伏关系[J]. 北京农业科学, 1990, 8(3): 10-13. ( 0) |
| [9] |
KASHIWAGI T, ISHIMARU K. Identification and functional analyses of a locus for improvement of lodging resistance in rice[J]. Plant Physiol, 2004, 134(2): 676-683. DOI:10.1104/pp.103.029355 ( 0) |
| [10] |
向达兵, 郭凯, 雷婷, 等. 磷钾营养对套作大豆茎秆形态和抗倒性的影响[J]. 中国油料作物学报, 2010, 32(3): 395-402. ( 0) |
| [11] |
谢甫练, 胡凤新, 赵庆祥, 等. 不同倒伏程度对大豆生育性状和产量性状的影响[J]. 辽宁农业科学, 1994(5): 43-45. ( 0) |
| [12] |
钟开珍, 梁江, 韦清源, 等. 大豆种质倒伏性遗传及其与主要农艺性状的相关分析[J]. 大豆科学, 2012, 31(5): 703-706. DOI:10.3969/j.issn.1000-9841.2012.05.003 ( 0) |
| [13] |
范冬梅, 杨振, 马占洲, 等. 多环境条件下大豆倒伏性相关形态性状的QTL分析[J]. 中国农业科学, 2012, 45(15): 3029-3039. ( 0) |
| [14] |
王松荣. 浅析大豆的播种过程及方法[J]. 科技创新与应用, 2013(20): 260. ( 0) |
| [15] |
HILTBRUNNER J, STREIT B, LIEDGENS M. Are seeding densities an opportunity to increase grain yield of winter wheat in a living mulch of white clover?[J]. Field Crop Res, 2007, 102: 163-171. DOI:10.1016/j.fcr.2007.03.009 ( 0) |
| [16] |
唐海涛, 田玉秀, 康继伟, 等. 玉米杂交种抗倒伏性的研究[J]. 种子, 2008, 27(11): 57-60. DOI:10.3969/j.issn.1001-4705.2008.11.017 ( 0) |
| [17] |
杨世民, 谢力, 郑顺林, 等. 氮肥水平和栽插密度对杂交稻茎秆理化特性与抗倒伏性的影响[J]. 作物学报, 2009, 35(1): 93-103. ( 0) |
| [18] |
谢甫绨, 曹海潮, 张惠君, 等. 大豆远缘杂交后代植株倒伏与农艺性状的关系[J]. 大豆科学, 2009, 28(5): 795-800. ( 0) |
| [19] |
王成雨, 代兴龙, 石玉华, 等. 氮肥水平和种植密度对冬小麦茎秆抗倒性能的影响[J]. 作物学报, 2012, 38(1): 121-128. ( 0) |
| [20] |
郑亭, 陈溢, 樊高琼, 等. 株行配置对带状条播小麦群体光环境及抗倒伏性能的影响[J]. 中国农业科学, 2013, 46(8): 1571-1582. DOI:10.3864/j.issn.0578-1752.2013.08.006 ( 0) |
| [21] |
陈小林, 杨文钰, 陈忠群, 等. 不同施氮水平下净、套作大豆茎秆特征比较研究[J]. 大豆科学, 2011, 30(1): 101-104. ( 0) |
| [22] |
刘唐兴, 官春云. 不同密度的油菜根系特征和产量与倒伏之间的相关性初探[J]. 西南农业学报, 2008, 21(1): 23-25. DOI:10.3969/j.issn.1001-4829.2008.01.005 ( 0) |
| [23] |
沈学善, 李金才, 屈会娟, 等. 砂姜黑土区小麦玉米秸秆全量还田对玉米抗倒性能的影响[J]. 中国农业科学, 2011, 44(10): 2005-2012. ( 0) |
| [24] |
李国辉, 钟旭华, 田卡, 等. 施氮对水稻茎秆抗倒伏能力的影响及其形态和力学机理[J]. 中国农业科学, 2013, 46(7): 1323-1334. DOI:10.3864/j.issn.0578-1752.2013.07.003 ( 0) |
| [25] |
徐磊, 王大伟, 时荣盛, 等. 小麦基部节间茎秆密度与抗倒性关系的研究[J]. 麦类作物学报, 2009, 29(4): 673-679. ( 0) |