 2015, Vol. 36
2015, Vol. 36



云南省大部分地区常年的降雨量都在1 100 mm左右,不属于干旱地区.但其年降雨量有着明显的季节性分布特点.每年雨季(5—10月)的降雨量占全年的80%~90%,且6—8月就达全年的60%左右.这种降水特点对当地的夏粮生产,尤其是对占该省小麦常年播种面积约2/3的旱地小麦(当地俗称地麦)的播种、出苗和灌浆成熟都极为不利.为保证地麦种子在播种后能正常出苗和生长,当地人通常将其播种期提前至雨季结束前的9月底至10月初.因此,在水分亏欠条件下小麦种子的萌发出苗能力,包括萌发期的抗旱力对保障田间基本苗至关重要.
大量的研究表明,不同小麦品种在渗透或干旱胁迫处理下种子萌发阶段的抗旱性与种子萌发率或发芽率、初生根和胚芽等生长、叶片各种酶的活性变化都有一定关系[1-7].在PEG溶液处理下的相对发芽率、芽鞘和主胚根长、侧根数、芽鞘和叶片脯氨酸、CAT和SOD含量的抗胁迫系数都可作为小麦种子萌发阶段抗旱性鉴定的指标[8-9].云南省有着丰富的地方小麦种质资源,关于云南旱地小麦抗旱性的研究已有一些报道[10-13],但鲜见关于不同种、亚种的全生育期抗旱性不同的小麦种质资源种子萌发期抗旱性相关性状的差异、及与全生育期抗旱性鉴定指标——抗旱指数、抗旱系数关系研究.因此,在前期经自然干旱胁迫、以抗旱指数为指标,筛选出2种5亚种48份全生育期抗旱性强和极强的品种[14]基础上,采用PEG高渗溶液摸拟干旱环境,研究了全生育期抗旱性不同的品种种子萌发期抗旱相关性状的差异,以期筛选到早期阶段能间接鉴定全生育期抗旱性的指标.
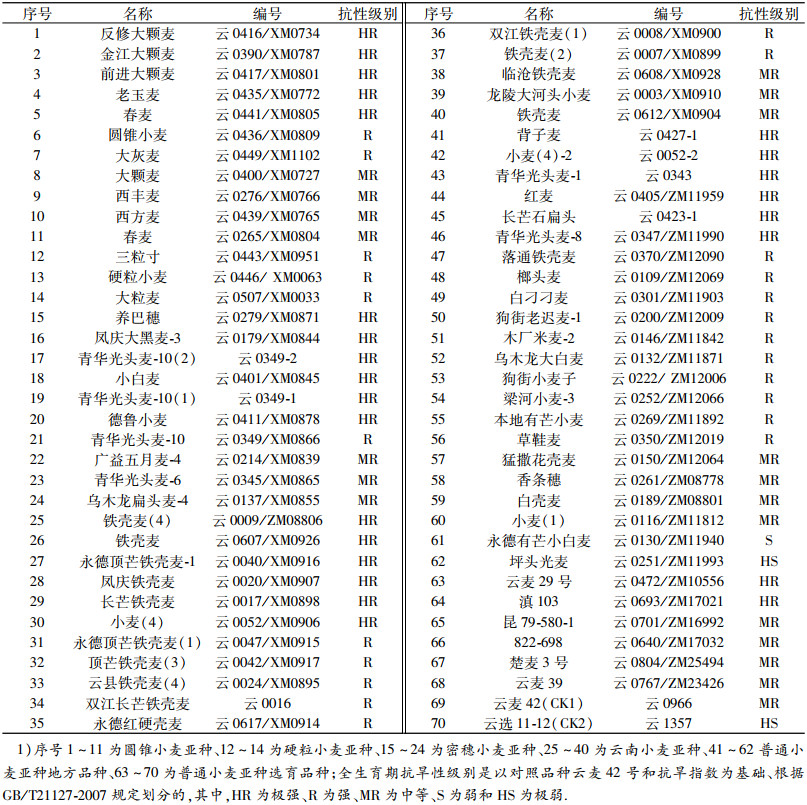
1 材料与方法 1.1 材料由云南省作物种质中期库保存和提供、经田间自然干旱胁迫鉴定表现为全生育期抗旱性不同的70份材料, 其中,全生育期抗旱性强-极强(R-HR)的材料48份,中等(MR)的19份和弱-极弱(S-HS)的3份(表 1).这些材料中包括了圆锥小麦Triticum turgidum ssp. turgidum亚种11份和硬粒小麦T. turgidum ssp. durum亚种3份,普通小麦T. aestivum ssp. aestivum亚种的地方品种22份和早期育成品种6份、密穗小麦T. aestivum ssp. compactum亚种10份和云南小麦T. aestivum ssp. yunnanense(俗称铁壳麦)亚种16份.全生育期抗旱性极弱(HS)的对照品种为云选11-12、中等的为云麦42号.
|
|
表 1 参试材料名称、编号及全生育期抗旱性级别1) Table 1 Names, codes and grades of drought resistance of tested materials in the whole growing period |
按照GB/T21127-2007[15]规定的要求,从每份参试材料中随机挑选出800粒无病虫的完整种子,以100粒为1个重复,分别装入事先贴有标签的8个培养皿,组成各重复4次的PEG胁迫和水培对照处理.然后,将15 mL去离子水和事先配制好的192 g·L-1的PEG-6000溶液分别加入对照和胁迫处理的各培养皿中,加盖.将培养皿放入(20±1) ℃的光照培养箱,按GB/T3543.4-1995[16]完成发芽试验.发芽试验前3 d不予种子光照,第4天开始8 h·d-1光照、补充适量水分或PEG溶液.
1.3 数据采集与统计分析在种子发芽至第4天和第8天时,按GB/T3543.3规定的标准分别统计培养72和168 h的种子发芽势和发芽率,并在第8天时随机记录和测量各处理及重复10株幼苗的胚根数、最大胚根长、胚芽鞘长和胚芽长4个性状的观测值.用Excel 2007分别计算出每份参试材料在2种处理下的性状相对值和根芽长度比.各性状的相对值=PEG处理下4次重复的平均值/水培处理下4次重复的平均值.其中,相对发芽势和发芽率以百分数(%)表示.利用IBM SPSS Statistics v19软件的单样本T测验分析全生育期抗旱性强-极强的品种的观测值及相对值与对照品种云选11-12的差异,用ANOVA模型分析全生育期抗旱性强-极强的品种的观测值、相对值与全生育期抗旱性中等、弱-极弱的品种间差异,用相关双变量模型分析全生育期抗旱性强-极强、中等的品种观测值及相对值与抗旱指数、抗旱系数的相关性.
2 结果与分析 2.1 全生育期抗旱性不同的品种种子萌发期抗旱相关性状的差异 2.1.1 不同亚种的全生育期抗旱性强-极强的品种与对照品种的种子萌发期抗旱性相关性状的差异与全生育期抗旱性极弱(HS)的对照品种云选11-12相比,在水培处理下,全生育期抗旱性强-极强(R-HR)的云南小麦和普通小麦选育品种的发芽势、硬粒小麦的发芽率、云南小麦和普通小麦地方品种的胚根数、密穗小麦和云南小麦的最大胚根长、云南小麦的胚芽长度、密穗小麦和云南小麦的根芽长比都显著或极显著地低于对照品种;圆锥小麦的胚根数以及圆锥小麦、密穗小麦、云南小麦和普通小麦地方品种的胚芽鞘长、密穗小麦的胚芽长则显著或极显著地高于对照品种.在PEG胁迫处理下,R-HR云南小麦的发芽势和发芽率、云南小麦和普通小麦地方品种的胚根数和最大胚根长、密穗小麦、云南小麦和普通小麦地方品种的根芽长度比也极显著地低于对照品种;圆锥小麦和普通小麦地方品种的发芽势、普通小麦选育品种的胚根数、圆锥小麦、密穗小麦、云南小麦和普通小麦地方品种的胚芽鞘长及云南小麦的胚芽长则显著或极显著地高于对照品种(表 2).
|
|
表 2 不同亚种的全生育期抗旱性强-极强的品种与极弱的对照品种云选11-12的种子萌发期抗旱性相关性状的平均值 Table 2 Average values of drought resistance traits of R-HR varieties and contrast "Yunxuan 11-12"of HS at seed germination stage |
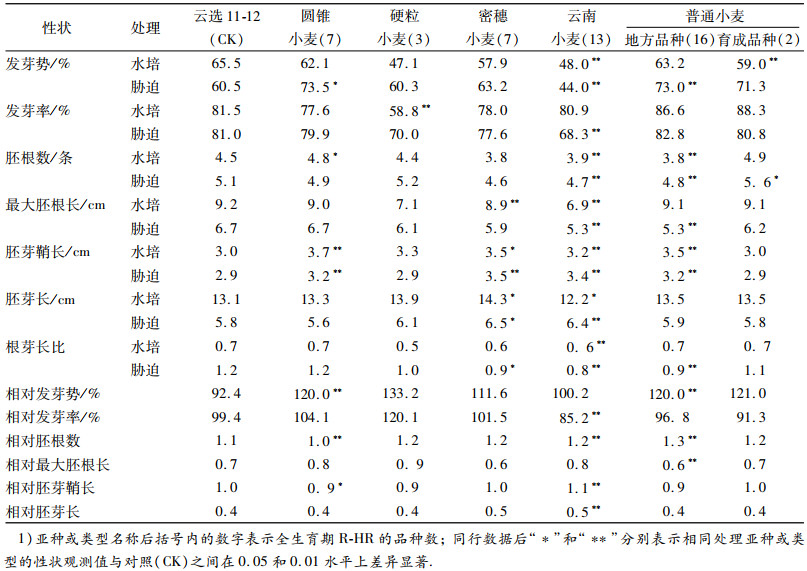
2种处理条件下,圆锥小麦和普通小麦地方品种的相对发芽势、云南小麦和普通小麦地方品种的相对胚根数、云南小麦的相对胚芽鞘长和相对胚芽长显著或极显著地高于对照品种;云南小麦的相对发芽率和普通小麦地方品种的相对最大胚根长、圆锥小麦的相对胚根数和相对胚芽鞘长则显著或极显著地低于对照品种.很显然,不同亚种的R-HR品种种子萌发期抗旱性相关性状与对照品种间的性状差异是不同的,在所有亚种的R-HR品种的种子萌发期抗旱性相关性状与对照品种的性状比较中,没有一个观察性状的平均值或相对值有与对照品种一致的变化趋势.相比之下,无论是在水培还是在PEG处理下,4个亚种的R-HR品种胚芽鞘长度都表现出了较对照品种更长的变化.说明不同亚种的R-HR品种种子萌发期性状受干旱影响的程度或对干旱环境的敏感性是不同的.因此,试图用同一个种子萌发期抗旱性相关性状完全把观察的所有亚种或类型的R-HR品种从试验样品中区分出来是不现实的.但是,用胚芽鞘长度能将圆锥小麦、密穗小麦、云南小麦亚种和普通小麦亚种地方品种的R-HR材料与极弱的选育对照品种云选11-12区分开.另外,在PEG处理下,用圆锥小麦亚种的发芽势和相对发芽势,云南小麦的胚芽长、相对胚根数、相对胚芽鞘长和相对胚芽长,普通小麦亚种地方品种的发芽势、相对发芽势和相对胚根数,选育品种的胚根数也分别能将这些亚种或类型的R-HR品种与极弱的对照品种区分开.
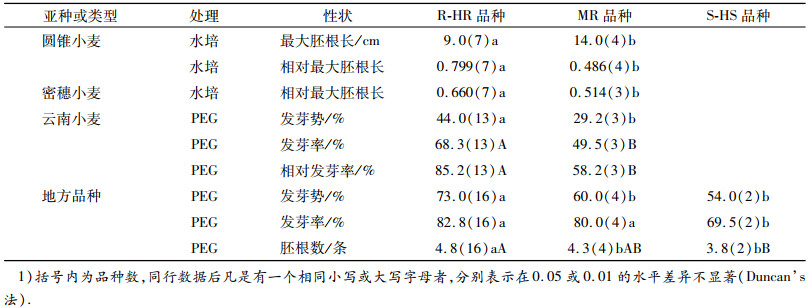
2.1.2 全生育期抗旱性强-极强的品种与中等、弱-极弱品种的种子萌发期抗旱性相关性状的差异与MR、S-HS品种相比,R-HR的品种中,除圆锥小麦在水培处理下的最大胚根长和相对最大胚根长、密穗小麦的相对最大胚根长、云南小麦在PEG胁迫处理下的发芽势、发芽率及相对发芽率、普通小麦地方品种在PEG胁迫处理下的发芽势、发芽率和胚根数,显著或极显著地较MR或S-HS品种的长或高外,其他观测值均无明显变化(表 3中未列出),同一性状在一些亚种的全生育期抗旱性不同的品种间有显著差异,而在另一些亚种中又无明显差异.5个亚种的R-HR品种与中等、弱-极弱品种的差异比较中,也没有一个性状的平均值或其相对值都有一致的显著性变化(表 3).这说明,利用全生育期抗旱性不同的品种间性状的差异不能完全将每一个亚种的R-HR品种从试验样品中挑选出来.利用相对最大胚根长能把圆锥小麦亚种和密穗小麦亚种、利用发芽势、发芽率及相对发芽率能把云南小麦亚种的R-HR品种与中等品种区分开,而利用发芽势和胚根数能把普通小麦地方品种R-HR材料与中等、弱-极弱的材料区分开.
|
|
表 3 不同亚种的全生育期抗旱性强-极强的品种与中等或弱-极弱的品种的种子萌发期抗旱性相关性状的差异1) Table 3 Difference of related drought resistance traits between R-HR and MR or S-HS varieties at seed germination stage |
在PEG处理下,R-HR品种抗旱指数(IDR)与圆锥小麦的根芽长度比(r=0.782*)、云南小麦亚种的相对胚根数(r=0.746**)、普通小麦地方品种的相对胚芽长(r=0.577*)呈显著或极显著的正相关,与圆锥小麦的胚根数(r=-0.756*)、硬粒小麦的相对最大胚根长(r=-0.997*)和密穗小麦的相对胚芽鞘长(r=-0.758*)呈显著负相关.在水培处理下,MR品种的IDR与密穗小麦的胚芽长(r=0.999*)呈显著正相关,与普通小麦选育品种的最大胚根长(r=-0.960*)、普通小麦地方品种的相对胚根数(r=-0.982*)呈显著负相关.说明虽然不是所有观察的种子萌发期抗旱相关性状都与不同亚种R-HR品种的IDR有显著相关,但是通过PEG胁迫处理并选择根芽长度比大、相对胚根数多和相对胚芽更长的材料,有望分别获得圆锥小麦、云南小麦和普通小麦地方品种R-HR育种材料,或者通过选择胚根数少、相对最大胚根更长和相对胚芽鞘更短而获得圆锥小麦、硬粒小麦和密穗小麦R-HR育种亲本.在PEG胁迫处理下选择相对胚根数少、或者在水培处理下选择胚芽更长和最大胚根长更短的材料, 有望获得普通小麦地方品种、密穗小麦和普通小麦品种的MR育种材料.
2.2.2 与抗旱系数的相关性在水培处理下,抗旱系数(CDR)与R-HR圆锥小麦的发芽率(r=-0.825*)、硬粒小麦的胚芽长(r=-0.999**)和密穗小麦的胚芽鞘长(r=0.966**)显著相关;与MR圆锥小麦的相对胚芽长(r=0.981*)和普通小麦地方品种的胚芽长(r=-0.963*)有显著相关.在PEG处理下,CDR与R-HR密穗小麦的根芽长度比(r=-0.788*)、普通小麦地方品种的最大胚根长(r=0.618*)、胚芽长(r=0.677**)和相对胚芽长(r=0.648**)、圆锥小麦的相对最大胚根长(r=0.798*)呈显著或极显著相关.表明在水培处理下,选择发芽率低、胚芽短、胚芽鞘较长、最大胚根和胚芽更长的材料,可能获得圆锥小麦、硬粒小麦和密穗小麦R-RH育种材料;在PEG处理下,选择根芽长度比小、最大胚根和胚芽更长、相对最大胚根和相对胚芽更长的材料,可能筛选到密穗小麦、普通小麦地方品种和圆锥小麦R-RH育种材料;选择相对胚芽较长的圆锥小麦或在水培处理下选择短胚芽的普通小麦地方品种,则可能获得MR育种材料.
3 讨论与结论小麦种子萌发或幼苗期相关性状,如胚根数、主胚根长、胚芽鞘长、胚芽长、根芽长度比及根冠比等都被认为是鉴定种子萌发或幼苗生长阶段抗旱性的指标.其中胚芽鞘长被认为与抗旱指数、抗旱系数有显著或极显著相关性[3-4, 9].在PEG胁迫处理下,抗旱性强的品种芽鞘长度显著较抗旱性弱的品种更长、生长更快[1-3].在本研究中,全生育期抗旱性强-极强(R-RH)的圆锥小麦、密穗小麦、云南小麦和普通小麦地方品种的胚芽鞘长度极显著地大于全生育期抗旱性极弱的对照品种云选11-12,这与前人[1-3]的研究结果一致.但R-RH品种与中等(MR)品种之间差异不显著,也就是说在PEG胁迫处理下,胚芽鞘的长度变化不能区分所有亚种R-RH与MR品种.可见,胚芽鞘长度对小麦抗旱性的指标作用是有限的.另外,由于作物抗旱性是受多基因控制的复杂数量性状[17],任何由单一的形态性状或指标鉴定出来的结果都可能是片面和不客观的,所以企图用胚芽鞘长度等单一性状来准确筛选出不同种、亚种的抗旱品种也是不现实的.
本研究结果表明,在PEG胁迫处理下,R-RH圆锥小麦的根芽比、云南铁壳麦的相对胚根数、普通小麦地方品种的相对胚芽长与抗旱指数有显著或极显著正相关,圆锥小麦的胚根数、硬粒小麦的相对最大胚根长、密穗小麦的相对胚芽鞘长与抗旱指数有显著负相关;圆锥小麦的相对最大胚根长、普通小麦地方品种的最大胚根长、胚芽长和相对胚芽长与抗旱系数有显著或极显著正相关,这与孙晓波[5]的研究结果类似.在PEG胁迫处理下,胚芽鞘长与抗旱指数、抗旱系数无显著相关;在水培处理下,MR密穗小麦的胚芽鞘长与抗旱系数有极显著正相关.本研究发现的与抗旱指数和抗旱系数有显著相关性的相对胚根数、相对最大胚根长、相对胚芽鞘长、相对胚芽长、在PEG胁迫处理下的胚根数和根芽长度比等性状,对不同亚种或类型的R-RH育种材料的早期间接筛选具有指导作用.由于抗旱指数包含了籽粒产量潜力,因此利用与抗旱指数显著相关的性状间接筛选出的抗旱材料可能更有育种利用价值.
| [1] |
王玮, 邹琦. 胚芽鞘长度作为冬小麦抗旱性鉴定指标的研究[J]. 作物学报, 1997, 23(4): 459-467. DOI:10.3321/j.issn:0496-3490.1997.04.012 (  0) 0) |
| [2] |
邹琦, 王玮, 杨兴洪, 等. 冬小麦抗旱性鉴定的新方法:低水势下胚芽鞘长度法[J]. 中国农学通报, 2000, 16(5): 23-25. DOI:10.3969/j.issn.1000-6850.2000.05.008 ( 0) |
| [3] |
关周博, 王士强, 陈亮, 等. 模拟干旱胁迫下冬小麦胚芽鞘长度变化及与抗旱性的关系研究[J]. 干旱地区农业研究, 2009, 27(4): 125-130. ( 0) |
| [4] |
王玮, 邹琦, 杨兴洪, 等. 水分胁迫下冬小麦芽鞘长度与抗旱性的关系及其遗传特性的研究[J]. 西北植物学报, 1998, 18(1): 24-29. DOI:10.3321/j.issn:1000-4025.1998.01.005 ( 0) |
| [5] |
孙晓波. 冬小麦抗旱性鉴定指标的研究[D]. 北京: 中国农业科学院, 2007: 11-29. http://kns.cnki.net/KCMS/detail/detail.aspx?filename=HBNB604.016&dbname=CJFD&dbcode=CJFQ
( 0) |
| [6] |
武仙山, 王正航, 昌小平, 等. 用株高旱胁迫系数分析小麦发育中的抗旱性动态[J]. 作物学报, 2008, 34(11): 2010-2018. ( 0) |
| [7] |
张玉梅, 林琪, 姜雯, 等. 渗透胁迫条件下不同抗旱性小麦品种萌发期生理生化指标的变化[J]. 麦类作物学报, 2006, 26(6): 125-130. DOI:10.3969/j.issn.1009-1041.2006.06.027 ( 0) |
| [8] |
景蕊莲, 昌小平. 用渗透胁迫鉴定小麦种子萌发期抗旱性的方法分析[J]. 植物遗传资源学报, 2003, 4(4): 292-296. DOI:10.3969/j.issn.1672-1810.2003.04.002 ( 0) |
| [9] |
张玉梅. 控水条件下小麦不同抗旱性鉴定指标的研究[D]. 青岛: 莱阳农学院, 2006: 13-56. http://www.wanfangdata.com.cn/details/detail.do?_type=degree&id=Y906097
( 0) |
| [10] |
程加省, 于亚雄, 勾宇宏, 等. 云南旱地小麦品种的抗旱性研究[J]. 云南农业大学学报, 2007, 22(1): 13-16. DOI:10.3969/j.issn.1004-390X.2007.01.003 ( 0) |
| [11] |
勾宇宏, 程加省, 杨金华, 等. 云南旱地小麦抗旱性的灰色关联分析[J]. 云南农业科技, 2007(4): 13-15. DOI:10.3969/j.issn.1000-0488.2007.04.004 ( 0) |
| [12] |
程加省, 于亚雄, 杨金华, 等. 旱地小麦品种抗旱性的生理生化指标分析及评价[J]. 大麦与谷类科学, 2007(3): 30-32. DOI:10.3969/j.issn.1673-6486.2007.03.011 ( 0) |
| [13] |
伍少云, 奉有壁. 云南小麦(Triticum aestivum ssp. yunnanense King)的抗旱性鉴定研究初报[J]. 植物遗传资源科学, 2001, 2(2): 27-30. DOI:10.3969/j.issn.1672-1810.2001.02.006 ( 0) |
| [14] |
周国雁, 伍少云. 不同云南小麦种质资源的全生育期抗旱性及与主要农艺性状的相关性[J]. 华南农业大学学报, 2013, 34(3): 309-314. ( 0) |
| [15] |
中华人民共和国国家质量监督检验检疫总局, 中国国家标准化管理委员会. GB/T21127-2007小麦抗旱性鉴定评价技术规范[S]. 北京: 中国标准出版社, 2008: 1-5.
( 0) |
| [16] |
国家技术监督局. GB/T3543. 4-1995农作物种子检验规程发芽试验[S]. 北京: 中国标准出版社, 1995: 1-20.
( 0) |
| [17] |
黄清华, 曹连莆, 张新忠, 等. 数量性状定位及作物抗旱性分析研究进展[J]. 安徽农业科学, 2007, 35(21): 6386-6388. DOI:10.3969/j.issn.0517-6611.2007.21.023 ( 0) |