 2015, Vol. 36
2015, Vol. 36



2. 东北师范大学 生命科学学院,吉林 长春 130024;
3. 吉林省前郭县农业局,吉林 松原 138000
2. School of Life Sciences, Northeast Normal University, Changchun 130024, China;
3. Agricultural Bureau of Qianguo County in Jilin Province, Songyuan 138000, China
1959年Coe在玉米田中发现了孤雌生殖诱导系Stock6,利用Stock6作父本与其他玉米材料进行杂交,可在其子代中获得一定数量的单倍体[1].基于该方法发展起来的单倍体育种技术[玉米DH(Doubled haploid)育种技术]与分子标记辅助育种技术、转基因技术形成了现代玉米育种技术的新体系[2].国内外各大公司、科研院所和高等院校相继开展了玉米DH育种,并用于实践[3].
采用常规技术获得一个配合力高、农艺性状优良的玉米自交系,需进行连续7~8代的人工自交与选择,并且耗时耗力[4].利用单倍体技术可大大缩短产生纯合品系(DH系)的时间,一般只需要2~3代,选育纯合材料的效率远远超过常规育种方法.但Stock6本身存在着诱导率偏低、花粉量偏少、抗病性差等严重缺陷[5].为此,国内外研究者对其进行了改良.Lashermes等[6]从Stock6×W23(ig)后代中选育出诱导系SW14;Geiger等[7]从KEMS×SW14杂交后代中选育出了RWS,诱导率为8.1%;Sarkar等[8]从Sock6回交后代中获得了诱导率超过5%的新材料;刘志增等[9]从Stock6×BHO(高油群体)后代中获得诱导率达5.34%的农大高诱1号;才卓等[10]以Stock6×M278为基础材料进行遗传改良,选育出诱导率在10%以上的吉高诱系3号.德国KWS公司、法国利马格兰公司和美国杜邦先锋公司选育出诱导率6%以上的诱导系,并已开始商业化应用[11].
玉米单倍体育种的首要条件是孤雌生殖诱导系的选育,加快育种进程要求高的单倍体诱导率.玉米单倍体诱导率的提高可以大大提高单倍体育种的效率,也可节约育种成本.Sarkar等[12]研究发现玉米单倍体的诱导率受诸多因素影响.本研究旨在分析和探讨玉米单倍体育种实践中影响诱导率的一些因素,进而提出相应的解决措施,为玉米单倍体育种实践提供参考.
1 材料与方法 1.1 母本基因型效应2011年夏选择10个自交系、10个杂交组合为诱导材料种植于吉林农业大学长春玉米育种基地,每个试验材料种植1行,行距65 cm,密度为6万株·hm-2,无区组重复,试验材料见表 1,基础材料涵盖了东北地区常用的自交系及主要的杂种优势模式.父本为JS6-1.
|
|
表 1 供试玉米自交系和杂交组合 Table 1 The tested maize inbred lines and hybridized combinations |
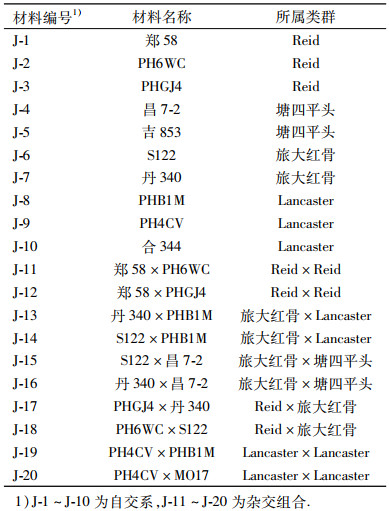
母本吐丝后授粉,杂交诱导10~20个果穗,收获后选择结实较好的果穗混合脱粒,进行单倍体的初步鉴定,确定拟单倍体(胚乳紫色且胚无色),所有拟单倍体于2012年春种植于吉林农业大学长春玉米育种基地进行田间鉴定,根据田间鉴定结果统计不同基础材料的单倍体诱导率.
1.2 不同种质的世代选择效应母本材料为吉林农业大学作物遗传教研室经过多轮选择不同世代的4大种质类群和热带种质群(表 2).父本为JS6-1,杂交诱导试验于2011年夏季在吉林农业大学长春玉米育种基地进行,秋季共获得不同世代的5个基础种质群250个杂交果穗48 642粒种子,其中拟单倍体2 806粒.2012年春对收获不同种质类群、不同世代的单倍体种子采用完全随机试验设计方法进行田间播种,单倍体鉴定方法同1.1.
|
|
表 2 供试母本材料 Table 2 The tested female parent materials |

试验原始数据先在Excel软件中进行处理,再用DPS[13]软件进行数据分析.计算群单倍体诱导率,方法同李国良等[14]的方法.
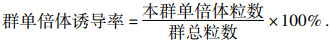
|
以JNY301、JNY611和JNY886(以上3品种为吉林农业大学玉米育种课题组选育品种)以及生产上种植面积较大的先玉335(XY335)和郑单958(ZD958)共5个杂交种作母本分别和诱导系JS6-2(农大高诱1号改良系)进行杂交诱导,试验材料见表 3.2011年在长春和三亚两地进行杂交诱导,从杂交组合中选择10个结实性好的果穗对单倍体诱导率进行测定.其中长春基地行距65 cm,密度为9万株·hm-2;三亚基地行距90 cm,密度为12万株·hm-2,单倍体的具体鉴定方法同1.1.
|
|
表 3 不同生态环境效应供试材料 Table 3 Experimental materials for testing under different ecological conditions |

2011年夏、2012年夏在吉林农业大学长春玉米育种基地分别以郑单958(ZD958)、JNY986、农华101(NH101)、JNY701为母本,以诱导系JS6-1为父本,选择不同的授粉时期进行授粉,根据母本吐丝后的天数划分授粉时期,即将吐丝时剪开苞叶授粉定义为0 d,刚吐丝时定义为1 d,依次往后记录吐丝后的天数,具体分为以下3个处理:早期授粉0~2 d;中期授粉3~5 d;晚期授粉6~8 d.单倍体的鉴定方法同1.1.
1.5 果穗不同部位的诱导率选择郑单958(ZD958)、先玉335(XY335)、利民33(LM33)、JNY986和农华101(NH101)5个杂交种于2011年冬天种植于吉林农业大学三亚育种基地,见表 4.以诱导系JS6-3(吉高诱3号改良系)作父本和各杂交种杂交30穗,收获后选择结实性好的果穗按底部和顶部分开脱粒(各占果穗长1/2),挑出拟单倍体,2012年夏在吉林长春进行单倍体的田间鉴定,具体鉴定方法同1.1.
|
|
表 4 果穗不同部位效应供试材料 Table 4 Experimental materials for testing different parts of the maize ear |

由表 5可知,诱导系JS6-1对各材料均具有诱导孤雌生殖单倍体的能力.不同母本基础材料经诱导后诱导率存在显著差异,诱导率最高的是J-18(PH6WC×S122),为10.13%,最低的是J-17(PHGJ4×丹340),为4.97%,平均诱导率为8.42%.
|
|
表 5 母本诱导基础材料的单倍体诱导率比较 Table 5 Comparisons of the haploid induced rates of different female parent materials |
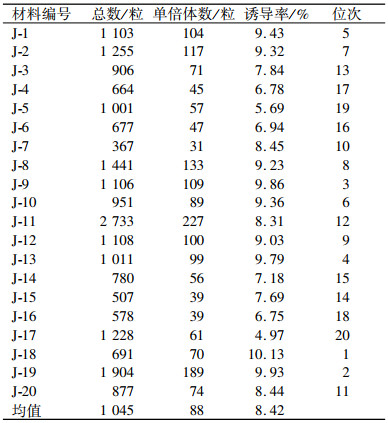
由表 6可知,在JS6-1诱导下,5个种质类群和5个种质世代均能产生单倍体,不同种质类群和种质世代的群单倍体诱导率的平均值差异明显,由此可以列出不同种质类群、不同种质世代单倍体诱导率的二因素方差分析表,见表 7.二因素方差分析表明:不同种质类群间P=0.006 4,达到极显著水平,不同世代间P=0.018,达到显著水平.
|
|
表 6 不同种质类群、不同世代的单倍体诱导率统计结果 Table 6 Statistical results of the haploid induced rates of different germplasms and generations |
|
|
表 7 不同种质类群、不同世代的单倍体诱导率方差分析表 Table 7 Variance analysis of the haploid induced rates of different germplasms and generations |
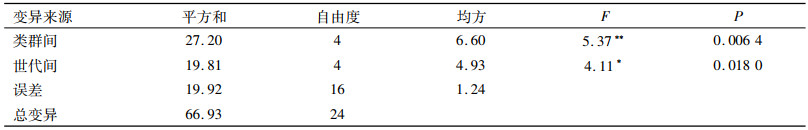
由表 8可知,在不同类群单倍体诱导率的多重比较中,诱导系JS6-1对Lancaster(4.94%)和Reid类群(4.88%)授粉所产生平均群单倍体诱导率比较高;对塘四平头和旅大红骨类群单倍体诱导率为4.16%和4.08%, 对热带种质群单倍体诱导率最低(2.12%).在玉米不同类群单倍体诱导率的多重比较中, JS6-1对4大类群授粉所产生的单倍体诱导率差异不显著, 其中对塘四平头和旅大红骨类群比热带处质群平均单倍体诱导率高, 差异显著(P < 0.05);对Lancaster和Reid类群授粉所产生平均群单倍体诱导率比热带种质平均单倍体诱导率高, 差异显著(P < 0.01).JS6-1对同一群体不同世代间诱导率表现为一般高世代S4、S5代单倍体诱导率高、每株单倍体粒数较低.S1、S2、S3代诱导率低、每株单倍体粒数较高, S1代单倍体诱导率较低, 但材料种穗大、结实率高, 实际得到的单倍体数量最多.JS6-1对S4、S5代平均群单倍体诱导率为5.14%, S4、S5代是S1~S3代平均单倍体诱导率(3.30%)的1.56倍.S4、S5代间单倍体诱导率差异不显著, S1~S3代间差异也不显著, 但S4、S5代与S1~S3代单倍体诱导率差异显著(P < 0.05).
|
|
表 8 不同种质群、不同世代间单倍体诱导率的多重比较1) Table 8 Multiple comparisons of the haploid induced rates of the different germplasm and generations |
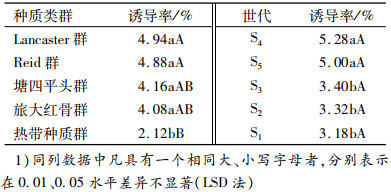
诱导系JS6-1对S1每株单倍体平均诱导数是12.36粒,比S4~S5平均每株单倍体诱导粒数(8.30粒)多4粒,即S1每株单倍体粒数显著高于S4~S5代(P < 0.05),S2~S3代每株单倍体粒数差异不显著,S4~S5代间亦如此.
2.2 生态环境、授粉时期和果穗部位对单倍体诱导率的影响三亚冬季单倍体诱导率高于长春夏季,其中XY335、ZD958、JNY611在三亚的诱导率(4.87%、4.35%、3.80%)显著高于长春的(3.95%、3.37%、2.09%)(P < 0.05),JNY886和JNY301在2个地点间的诱导率差异不显著.三亚的单倍体诱导率极显著高于长春(P < 0.01),其中三亚冬季的平均诱导率为3.15%,而长春夏季的诱导率仅为1.99%(表 9).
|
|
表 9 2010—2013年长春和三亚两地点间的诱导率比较 Table 9 Comparisons of the haploid induced rates between Changchun and Sanya from 2010 to 2013 |

由表 10可知,延迟授粉一般有降低玉米单倍体诱导率的趋势,除2011年以JNY701为母本,2012年以ZD958、JNY701为母本,晚期授粉时的单倍体诱导率高于中期授粉的诱导率.此外,早期授粉的单倍体平均诱导率最高(1.99%),其次是中期授粉(1.87%),晚期授粉的平均诱导率最低(1.59%).
|
|
表 10 不同母本诱导基础吐丝后不同天数和JS6-1杂交后的单倍体频率比较 Table 10 Comparisons of the haploid induced rates under different pollination stages after silking |

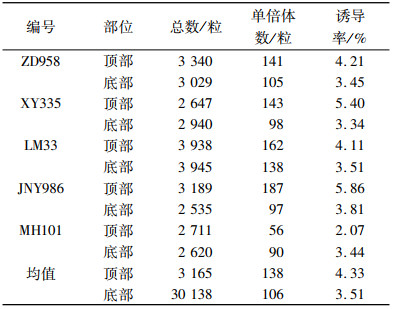
由表 11可知,除NH101之外,其余4个杂交果穗中顶部出现的单倍体频率均高于底部,其中顶部单倍体平均诱导率为4.33%,底部的为3.51%.
|
|
表 11 果穗顶部和底部的诱导率比较 Table 11 Comparisons of the haploid induced rates between top and bottom parts of ear |
本研究表明单倍体诱导率高低与母本基因型有关,彼此间差异显著,这与Rober等[15]、刘志增等[9]等的研究结果相一致.Chase[16]认为这种影响是由于母本基因型所产生的雌配子中有害基因的频率不同造成的.本研究结果表明Lancaster、Reid类群材料的单倍体诱导率显著高于塘四平头类群、旅大红骨类群、地方种质群和热带种质群.
3.2 诱导世代的选择Lashermes等[17]以Stock6作父本,以169wx/Syn421等3个组合作母本,比较了F2及F3世代的单倍体诱导率,得出两世代间的相关性只有0.59.李国良等[14]研究得出早代S1为最佳的单倍体诱导世代.本研究表明也得出了类似结论:在不同类群的高世代中S4~S5单倍体诱导率高于S1~S3,但每株获得的单倍体粒数却低于后者;S1单倍体诱导率较低,但收获的材料果穗大,实际得到的单倍体数量最多.S1是植株性状急剧分离的世代,可以通过植株抗病性、株高等田间农艺性状去除一些伪单倍体植株,选择一些比较优良的单株进行诱导获得单倍体,可以大大减少工作量,提高优良DH系出现的概率,因此本研究认为早代S1为单倍体最佳诱导世代.
综合考虑,如果双亲差异大,后代分离的基因较多,则仅靠1次重组就需要很大的群体才可能获得优良重组基因型.从玉米育种角度来说建议此类材料可以在S1代或更高世代诱导,而对于窄基群体则可以在S0代诱导.
3.3 诱导环境的优化Prigge等[17]在热带地区进行单倍体诱导得出2个环境下单倍体的诱导率具有显著差异,但Eder等[18]对12份基础材料在2种不同的生态环境下诱导并没有发现单倍体诱导率存在显著差异.黎亮等[19]多年试验结果也证实了在海南冬季进行单倍体诱导的效率显著高于北京夏季.本研究发现三亚冬季的诱导率显著高于长春夏季.长春夏季气温较高,三亚冬季气温较低且昼夜温差大,而气温较低的情况下可能更适合籽粒发育及色素标记基因的表达.
3.4 诱导时期的优化Chase[16]和Seaney等[20]的研究结果都表明延迟授粉可以提高玉米单倍体诱导率.本研究发现果穗顶部籽粒的单倍体诱导率高于底部,这与Sarkar等[12]、Chalyk等[21]的研究结果相似.这可能与卵细胞成熟的不同步、营养功能的变化、某些外界因子的局部影响等有关.本研究发现早期授粉的单倍体诱导率高于晚期授粉,此现象尚无法用花丝长短进行解释.Rotarenco等[22]认为这与异雄核受精有关,由于吐丝时间长的花丝直径更大,可以容纳更多的花粉管生长,单受精的概率可能降低,进而影响单倍体出现的概率.不同生态环境下单倍体诱导率的差异也可能与花丝自身结构以及花粉的活力有关.
| [1] |
邢锦丰, 张如养, 段民孝, 等. 单倍体技术在玉米育种中的应用及其问题探讨[J]. 作物杂志, 2012(2): 15-17. DOI:10.3969/j.issn.1001-7283.2012.02.004 (  0) 0) |
| [2] |
张如养, 段民孝, 赵久然, 等. 单倍体技术在玉米种质改良和育种中的应用方向[J]. 作物杂志, 2012(5): 4-7. ( 0) |
| [3] |
姜龙, 慈佳宾, 崔学宇, 等. 不同生态条件下玉米单倍体诱导率和加倍率研究[J]. 吉林农业大学学报, 2014, 36(2): 139-143. ( 0) |
| [4] |
刘纪麟. 玉米育种学[M]. 2版. 北京: 中国农业出版社, 2002: 172-177.
( 0) |
| [5] |
CHANG M T. Stock6 induced double haploidy is random[J]. Maize Genet Coop Newslett, 1992, 67: 98-99. ( 0) |
| [6] |
LASHERMES P, BECKERT M. Genetic control of maternal haploid in maize(Zea mays L.) and selection of haploid inducing lines[J]. Theor Appl Genet, 1988, 76: 405-410. ( 0) |
| [7] |
GEIGER H H, GORDILLO G A. Doubled haploids in hybrid maize breeding[J]. Maydica, 2009, 54: 485-499. ( 0) |
| [8] |
SARKAR K R, SUDHA P. Development of maternal-haploidy-inducer lines in maize[J]. Indian J Agri Sci, 1972, 42: 81-786. ( 0) |
| [9] |
刘志增, 宋同明. 玉米高频率孤雌生殖单倍体诱导系的选育与鉴定[J]. 作物学报, 2000, 26(5): 570-574. DOI:10.3321/j.issn:0496-3490.2000.05.009 ( 0) |
| [10] |
才卓, 徐国良, 刘向辉, 等. 玉米高频率单倍体生殖诱导系吉高诱系3号的选育[J]. 玉米科学, 2007, 12(1): 1-4. DOI:10.3969/j.issn.1005-0906.2007.01.001 ( 0) |
| [11] |
贺正华, 高立, 张士龙, 等. 玉米单倍体诱导系选育及诱导效果[J]. 湖北农业科学, 2014, 53(19): 4533-4536. ( 0) |
| [12] |
SARKAR K R, PRASANNA B M, GAYEN P. Distribution of haploids on the ear[J]. Maize Genet Coop Newslett, 1995, 69: 107. ( 0) |
| [13] |
唐启义, 冯明光. 实用统计分析及其DPS数据处理系统[M]. 北京: 科学出版社, 2002: 304-311.
( 0) |
| [14] |
李国良, 苏俊, 李春霞, 等. 农大高诱1号对玉米不同种质和世代单倍体诱导频率的研究[J]. 玉米科学, 2008, 16(5): 3-6. ( 0) |
| [15] |
ROEBER F K, GORDILLO G A, GEIGER H H. In vivo haploid induction in maize-performance of new inducers and significance of doubled haploid lines in hybrid breeding[J]. Maydica, 2005, 50(3/4): 275-283. ( 0) |
| [16] |
CHASE S S. Monoploids and monoploid derivatives of maize (Zea mays L.)[J]. Bot Review, 1969, 35: 117-167. DOI:10.1007/BF02858912 ( 0) |
| [17] |
PRIGGE V, ANCHEZ C, DHILLON B S, et al. Doubled haploids in tropical maize:Ⅰ:Effects of inducers and source germplasm on in-vivo haploid induction rates[J]. Crop Sci, 2011, 51(4): 149-150. ( 0) |
| [18] |
EDER J, CHALYK S. In vivo haploid induction in maize[J]. Theor Appl Genet, 2002, 104(4): 703-708. DOI:10.1007/s00122-001-0773-4 ( 0) |
| [19] |
黎亮, 李浩川, 徐小炜, 等. 玉米孤雌生殖单倍体诱导效率优化方法研究[J]. 中国农业大学学报, 2012, 17(1): 9-13. ( 0) |
| [20] |
SEANEY R R. Monoploids in maize[J]. Maize Genet Coop Newslett, 1954, 28: 22. ( 0) |
| [21] |
CHALYK S T. Creting new haploid-inducing lines of maize[J]. Maize Genet Coop Newslett, 1999, 73: 53-54. ( 0) |
| [22] |
ROTARENCO V A, MIHAILOV M E. The influence of ear age on the frequency of maternal haploids produced by a haploid-inducing lines[J]. Maize Genet Coop Newslett, 2007, 81: 9-10. ( 0) |