 2015, Vol. 36
2015, Vol. 36



随着经济发展、人口增长和生活水平的提高,能源危机、粮食短缺、环境污染等问题日益严重,已引起世界各国的广泛关注[1-2].为了逐渐摆脱对化石能源的过度依赖,保证粮食安全,减少温室气体排放等,各国纷纷研究适合自己国情的生物质能源.基于社会的可持续发展,以能源植物为主发展生物质能源是较理想的选择[3-4].能源植物具有光合效率高、生物质产量高、抗逆性强、生态效应好、生产成本低和易推广等特点,除可直接燃烧产生热能外,还可转化成常规的固态、液态和气态燃料[5].其中草本能源植物还因生长迅速、生长周期短和资源丰富等特点得到了广泛利用[6],如芒草Miscanthus spp.、王草Pennisetum purpureum×P. americanum、柳枝稷Panicum virgatum、芦竹Arundo donax、虉草Phalaris arundinacea等作为生物能源的主要原料,具有广阔的开发利用前景.
‘热研4号’王草Pennisetum purpureum×P. americanum cv. Reyan No.4是禾本科狼尾草属高秆多年生草本,原产于热带非洲,是象草P. purpureum和美洲狼尾草P. americanum的杂交种,以优质、高产而著称,被誉为“草中之王”[7].1984年中国热带牧草研究中心从哥伦比亚引种到海南岛,试种成功后逐步在全国各地推广.五节芒Miscanthus floridulus属禾本科芒属,主要分布东亚至波利尼西亚及澳大利亚以东,在我国主要分布在亚热带地区.2种草均具有生长迅速、生物量高、利用年限长和适应性强等特点[8-10].现已在我国广东、广西、福建、四川、江西等地广泛栽培[11-12].目前,对‘热研4号’王草的研究多集中在种植管理、转化工艺等方面,对五节芒多集中于分布、资源量、遗传多样性等方面.本文通过研究‘热研4号’王草和五节芒的光合特性日变化及其与环境因子间的相关关系,以期为‘热研4号’王草和五节芒的种植管理和利用推广提供依据.
1 材料与方法 1.1 试验地概况试验地位于华南农业大学增城宁西试验基地(N 23°14′,E 113°38′),属亚热带季风气候.年平均气温21.6 ℃,最热月(7月)的平均气温29.4 ℃,极端最高气温38.6 ℃;最冷月(1月)的平均气温13.3 ℃,极端最低气温-1.9 ℃.全年积温7 910.9 ℃,年均降水量为1 967.8 mm,年均太阳辐射值为4 367.2~4 597.3 MJ·m-2,年均日照1 707.2 h.
1.2 试验材料‘热研4号’王草和五节芒分别来自中国热带农业科学院热带作物品种资源研究所和中国科学院华南植物园.于2013年3月4日种植,每种草设3个试验小区,每小区面积14 m2(3.5 m×4.0 m),小区间距100 cm.‘热研4号’王草采用粗壮、无病害的茎芽作种茎,株行距60 cm;五节芒采用具有萌发能力的根茎作种茎,株行距80 cm.种植后于4月8日施肥、浇水,施肥量每小区均为622 g(N-P2O5-K2O有效养分质量分数为36%).
1.3 测定方法试验于2013年5月13日、7月13日、9月2日采用LI-6400便携式光合作用测定仪(Li-Cor, USA)对2种草的光合特性指标进行测定,包括气温(θa)、叶温(θl)、空气相对湿度(RH)、大气CO2摩尔分数(xa)、净光合速率(Pn)、蒸腾速率(Tr)、气孔导度(Gs)、胞间CO2摩尔分数(xi).测定过程中,设定叶室温度为20 ℃,空气流量500 μmol·s-1,光合有效辐射(PAR)1 000 μmol·m-2·s-1.测定时选取充分展开的健康叶片中部,每片每次连续读取3个稳定的数据,共3次重复.测定时间为09:00—17:00,每隔2 h测定1次.根据测定的指标计算叶片气孔限制值(Ls)、水分利用效率(WUE)和光能利用率(LUE),计算公式如下:
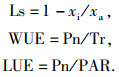
|
采用Excel 2007和SPSS 17.0软件对试验数据进行统计分析.
2 结果与分析2种草3次(5月13日、7月13日、9月2日)测定的光合特性日变化趋势较一致,图 1和图 2为5月份的测出结果.

|
图 1 环境因子日变化 Figure 1 Diurnal variation of environmental factors |

|
图 2 ‘热研4号’王草和五节芒的光合特性日变化 Figure 2 Diurnal variation in photosynthetic characteristics of Pennisetum purpureum×P. americanum cv. Reyan No.4 and Miscanthus floridulus |
植物的光合作用受多种环境因子的影响,其中温度、空气相对湿度和CO2摩尔分数是影响光合作用的主要环境因素[13].环境因子日变化如图 1所示,θa日变化呈单峰曲线,全天平均29.06 ℃,从09:00时开始逐渐升高,在15:00时左右达到全天最高温度(31.16 ℃),然后迅速下降.RH在09:00时左右达到最高(65.39%),随着时间的推移,因θa逐渐升高,导致RH逐渐降低.xa变化趋势呈倒钟型,09:00时左右开始,随着植物光合作用的进行,xa缓慢降低,午后逐渐回升;13:00时左右出现最低值(356.87 μmol·mol-1),17:00时左右出现全天最高值(482.21 μmol·mol-1),表现为早晚高、中午低,其原因可能是植物在早晚呼吸作用较强,而中午光合作用较强.
2.2 光合特性日变化 2.2.1 叶温、净光合速率、光能利用效率2种草的θl与θa日变化趋势相同.从图 2 a可见,θl从09:00至15:00不断升高达到最大值后,开始下降.Pn的日变化曲线(图 2 b)较为一致,呈单峰型,说明均无光合“午休”现象;从09:00开始随着θl的升高而逐渐升高,在13:00左右达到全天最大值,‘热研4号’王草和五节芒分别为31.97和29.08 μmol·m-2·s-1,此后逐渐下降;‘热研4号’王草的日均Pn为25.45 μmol·m-2·s-1,略高于五节芒(25.08 μmol · m-2·s-1). LUE日变化趋势(图 2c)与Pn相同,从09:00至13:00不断升高,达到最大值后逐渐下降,其中‘热研4号’王草的最大值(3.2%)高于五节芒(2.9%),说明‘热研4号’王草的光能利用效率高于五节芒.
2.2.2 蒸腾速率、水分利用效率从图 2 d、2 b可见,‘热研4号’王草和五节芒的Tr与Pn变化趋势相同,均在13:00时左右达到最大,分别为5.68和4.70 mmol·m-2·s-1,此时2种牧草的蒸腾作用最强,失水最快,其原因为θl从09:00至15:00逐渐升高,叶片内外细胞蒸气压增大,所以蒸腾速率加快.从图 2e看出,‘热研4号’王草和五节芒的WUE日变化均从09:00至17:00逐渐降低,可能是由于θa不断升高,Tr的加快而造成WUE逐渐下降,其日均值分别为6.06和6.44 μmol·mmol-1.
2.2.3 气孔导度、气孔限制值、胞间CO2摩尔分数在图 2 f中,‘热研4号’王草和五节芒的Gs日变化趋势较为一致,09:00和13:00均较高,最大值出现在13:00左右,分别为0.21和0.19 μmol·m-2·s-1,然后逐渐下降,在17:00左右时最低. Ls与xi日变化趋势正好相反,Ls日变化呈正钟型,均在11:00左右达最高值(均为0.86 μmol·m-2·s-1),早晚最低(图 2 g).xi日变化呈倒钟型,均在11:00左右最低,早晚最高,‘热研4号’王草和五节芒在09:00左右时xi分别为182.95和121.05 μmol·mol-1,在17:00左右时分别为202.13和220.13 μmol·mol-1(图 2 h).
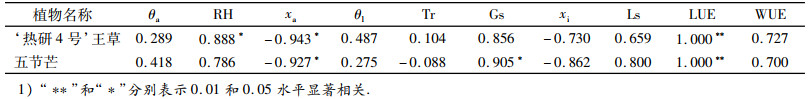
2.3 ‘热研4号’王草和五节芒净光合速率与主要影响因子的相关性分析2种草净光合速率与主要影响因子的相关关系如表 1.因测定时PAR设定为1 000 μmol·m-2·s-1,所以‘热研4号’王草和五节芒的Pn均与LUE(LUE=Pn/PAR)呈极显著正相关(P<0.01,r=1.000).此外,‘热研4号’王草Pn与RH呈显著正相关(P<0.05,r=0.888),与xa呈显著负相关(P<0.05,r=-0.943);五节芒Pn与Gs呈显著正相关(P<0.05,r=0.905),与xa呈显著负相关(P<0.05,r=-0.927).
|
|
表 1 2种草净光合速率与主要影响因子的相关性分析1) Table 1 Correlation analyses between net photosynthetic rate and major factors of two grasses |
植物的光合作用日变化是其物质积累和代谢的基本单元,也是分析环境因素影响植物生长和代谢的重要手段[14-15],它反映了植物全天光合作用随环境而发生的变化,是植物对特定环境条件适应的结果.温度通过净光合速率而影响光合作用,随着温度(10~35 ℃)的升高,作用于光合作用的酶活性不断提高,进而提高净光合速率[16].其中,气温主要是通过改变叶温影响植物的光合作用.本试验的气温和叶温在26~31 ℃,所以对提高净光合速率有促进作用.此外,温度不断升高使大气和植物叶片附近的相对湿度逐渐降低,从而加速蒸腾过程,然而蒸腾具有调节植物体温度的功能,所以可减缓热害[17].因此,随着温度的升高,植物光合作用增强是午间大气相对湿度和CO2摩尔分数降低的重要原因.
3.2 光合特性日变化 3.2.1 净光合速率、光能利用效率净光合速率是反映植物光合作用强弱的最重要指标,净光合速率越大,表明植物的光合作用越强.植物的净光合速率日变化曲线多呈单峰型或双峰型[18-19].C4植物在高温或干旱时,可利用细胞间隙低浓度的CO2进行光合作用,所以其净光合速率不存在光合“午休”现象[20].光能利用效率能反映植物对弱光的利用能力[21].‘热研4号’王草和五节芒的净光合速率和光能利用效率均呈单峰型,无光合“午休”现象,且‘热研4号’王草(31.97 μmol·m-2·s-1、3.2%)的最高值高于五节芒(29.08 μmol·m-2·s-1、2.9%),说明‘热研4号’王草的光合能力比五节芒强.种植半年后(9月5日)的干草产量‘热研4号’王草(22.12 t·hm-2)比五节芒(12.68 t·hm-2)高74.45%,说明确实是‘热研4号’王草的生产能力比五节芒高.
3.2.2 蒸腾速率、水分利用效率蒸腾作用是反映植物体内水分代谢状况或植物对水分利用效率的重要生理指标,植物在进行蒸腾作用时叶片气孔开放,有利于光合产物的形成[22-23].因植物在进行光合作用时需要不断供应水分及通过水分运载矿质营养成分,所以一般植物的光合速率越高,蒸腾速率越高[24-25].2种草的蒸腾速率与净光合速率相同,均在13:00时左右达到最大,且‘热研4号’王草高于五节芒,说明在高温下‘热研4号’王草比五节芒运输水分、矿物质和固定CO2的能力强.
水分利用效率由植物叶片的净光合速率和蒸腾速率共同决定[26-28],反映了植物光合作用和对环境的适应能力[20],是衡量植物消耗水分与生产物质之间关系的重要综合指标[29-30].‘热研4号’王草和五节芒的水分利用效率日均分别为6.04和6.44 μmol·mmol-1,说明‘热研4号’王草的水分利用效率略低于五节芒.
3.2.3 气孔导度、气孔限制值、CO2摩尔分数气孔是植物叶片与外界进行气体交换的主要通道,其闭合程度直接影响光合作用和蒸腾作用,进而减少植物叶片水分的过度散失,保证了水分的有效利用[19, 31],气孔导度则是反映这种交换能力的重要指标[32].‘热研4号’王草和五节芒的气孔导度在13:00时左右最大,且‘热研4号’王草高于五节芒,说明此时‘热研4号’王草叶片的气孔张开程度高于五节芒,其交换能力强于五节芒,从而有利于CO2的同化.因气孔导度的最大值出现在13:00时左右,而气温和叶温全天最高在15:00时左右,说明为防止植株叶片水分的过度散失,在达到全天最高温度时,2种草的气孔已提前收缩.
气孔限制值反映了植物叶片对大气CO2的相对利用率[19].2种草的气孔限制值均在11:00时左右达最高值,早晚低;胞间CO2摩尔分数日变化趋势与之相反,在17:00时左右最高,而此时2种草的净光合速率最低,说明植物的呼吸作用占主导地位,CO2开始在细胞内和细胞间积累,导致其浓度上升.
CO2是光合作用的主要原料,在农业生产中使用CO2气体施肥可明显提高产量[33].本试验中随着净光合速率的提高,大气和胞间CO2摩尔分数呈降低趋势,当净光合速率最低时,大气和胞间CO2摩尔分数迅速上升.说明当净光合速率较快时,植物进行光合作用固定的CO2较多,所以引起CO2摩尔分数下降.当CO2摩尔分数最低时,便会抑制净光合速率的提高.
3.3 2种草净光合速率与主要影响因子的相关性分析植物在外界环境因子的影响下,θl、Tr、xi、Gs等光合特性指标发生变化,最终引起Pn变化.光合作用是复杂的生理过程,其净光合速率日变化不仅由其自身生理特征决定,而且还与外界环境条件的变化关系密切[34].‘热研4号’王草和五节芒Pn均与xa呈显著负相关,且相关性最大,其次分别与RH和Gs呈显著正相关(P<0.05).所以, 影响‘热研4号’王草净光合速率的主要因子是大气CO2摩尔分数和空气相对湿度,影响五节芒的主要是大气CO2摩尔分数和气孔导度.
近年,我国面临着耕地面积不断减少、生态环境整体恶化、水资源短缺的严峻挑战,资源条件对农业发展的约束更加突出.因此,应充分利用农业废弃地、农田边际土地及冬季农闲田等土地资源,大力合理种植优良能源草,既能缓解煤炭、石油的供应压力、降低环境污染,又可有效改良土壤结构,避免与粮争地造成的粮食危机.本文对潜力较大的‘热研4号’王草和五节芒的光合特性的研究结果,可为相应地区的草种选择和合理的栽培管理提供参考依据.
3.4 结论‘热研4号’王草和五节芒的净光合速率日变化均呈“单峰型”,无光合“午休”现象,且‘热研4号’王草略高于五节芒.蒸腾速率、水分利用效率、光能利用率和气孔导度均在13:00时左右达最大值,除水分利用效率外,其余均为‘热研4号’王草略高于五节芒.气孔限制值日变化呈正钟型,早晚最低,胞间CO2摩尔分数日变化趋势则与之相反.影响‘热研4号’王草净光合速率的主要因子是大气CO2摩尔分数和空气相对湿度,影响五节芒的主要是大气CO2摩尔分数和气孔导度.总体来看,‘热研4号’王草的光合能力比五节芒强.
致谢:感谢中国科学院华南植物园宁祖林博士为本研究提供五节芒材料!
| [1] |
于建仁, 张曾, 迟聪聪. 生物质精炼与制浆造纸工业相结合的研究[J]. 中国造纸学报, 2008, 23(1): 80-84. DOI:10.3321/j.issn:1000-6842.2008.01.019 (  0) 0) |
| [2] |
NAGLE N, IBSEN K, JENNINGS E. A process economic approach to develop a dilute-acid cellulose hydrolysis process to produce ethanol from biomass[J]. Appl Biochem Biotechnol, 1999(77/78/79): 599-607. ( 0) |
| [3] |
吴国江, 刘杰, 娄治平, 等. 能源植物的研究现状及发展建议[J]. 科技与社会, 2006, 21(1): 53-57. ( 0) |
| [4] |
秦向华. 能源植物姗姗走来[J]. 森林与人类, 2004, 24(11): 49. ( 0) |
| [5] |
郭廷杰. 贯彻科学发展观拓展资源再生产业促进生物质能利用[J]. 再生资源研究, 2005(2): 1-4. DOI:10.3969/j.issn.1674-0912.2005.02.001 ( 0) |
| [6] |
高瑞芳, 张建国. 能源草研究进展[J]. 草原与草坪, 2013, 33(1): 89-96. DOI:10.3969/j.issn.1009-5500.2013.01.018 ( 0) |
| [7] |
孙凡. 草中之王:皇竹草[J]. 农技服务, 2004(9): 21-22. DOI:10.3969/j.issn.1004-8421.2004.09.013 ( 0) |
| [8] |
刘国道. 热带牧草栽培学[M]. 北京: 中国农业出版社, 2006: 66-67.
( 0) |
| [9] |
PENG Kejian, LI Xiangdong, LUO Chunling, et al. Vegetation composition and heavy metal uptake by wild plants at three contaminated sites in Xiangxi area, China[J]. J Environ Sci Heal A, 2006, 41(1): 65-76. DOI:10.1080/10934520500298838 ( 0) |
| [10] |
SCEBBA F, ARDUINI I, ERCOLI L, et al. Cadmium effects on growth and antioxidant enzymes activities in Miscanthus sinensis[J]. Biol Plantarum, 2006, 50(4): 688-692. DOI:10.1007/s10535-006-0107-0 ( 0) |
| [11] |
解新明. 草资源学[M]. 广州: 华南理工大学出版社, 2009: 130.
( 0) |
| [12] |
邓念丹, 刘清波, 蒋建雄, 等. 五节芒核心种质的构建研究概述[J]. 作物研究, 2010, 24(1): 130-134. ( 0) |
| [13] |
程鹏, 马晓红, 沈强, 等. 2个水仙新品种的光合作用特性研究[J]. 上海交通大学学报:农业科学版, 2013, 31(1): 28-33, 94. ( 0) |
| [14] |
冯建灿, 张玉洁. 喜树光合速率日变化及其影响因子的研究[J]. 林业科学, 2002, 38(4): 34-39. DOI:10.3321/j.issn:1001-7488.2002.04.006 ( 0) |
| [15] |
王忠. 植物生理学[M]. 北京: 中国农业出版社, 2000.
( 0) |
| [16] |
赵宗方, 凌裕平, 吴建华, 等. 梨树的光合特性[J]. 果树科学, 1993, 10(3): 154-156. ( 0) |
| [17] |
潘瑞枳. 植物生理学[M]. 2版. 北京: 高等教育出版社, 1982: 101-111.
( 0) |
| [18] |
李建栋, 刘岩, 高春雨, 等. 苦碟子光合作用日变化及其与环境因子关系的研究[J]. 人参研究, 2008, 20(2): 12-15. DOI:10.3969/j.issn.1671-1521.2008.02.003 ( 0) |
| [19] |
FARQUHAR G D, SHARKEY T D. Stomatal conductance and photosynthesis[J]. Ann Rev Plant Physiol, 1982, 33(1): 317-345. DOI:10.1146/annurev.pp.33.060182.001533 ( 0) |
| [20] |
许大全, 李德耀, 沈允钢, 等. 田间小麦叶片光合作用"午睡"现象的研究[J]. 植物生理学报, 1984, 10(3): 269-276. ( 0) |
| [21] |
许大全, 徐宝基, 沈允钢. C3植物光合效率的日变化[J]. 植物生理学报, 1990, 16(1): 1-5. ( 0) |
| [22] |
李育花, 任坚毅, 林玥, 等. 独叶草的光合生理生态特性[J]. 生态学杂志, 2007, 26(7): 1038-1042. DOI:10.3321/j.issn:1000-4890.2007.07.013 ( 0) |
| [23] |
刘金祥, 陈睿俊. 香根草、黑麦草和高丹草光合生理生态日变化的比较研究[J]. 热带农业科学, 2008, 28(4): 32-37. DOI:10.3969/j.issn.1009-2196.2008.04.007 ( 0) |
| [24] |
刘金祥, 陈伟云, 肖生鸿. 黑籽雀稗的光合生理特性研究[J]. 草业学报, 2009, 18(6): 254-258. DOI:10.3321/j.issn:1004-5759.2009.06.035 ( 0) |
| [25] |
孙启忠, 桂荣, 那日苏, 等. 赤峰地区不同生长年限沙打旺生产力的研究[J]. 中国草地, 1999(5): 30-35. ( 0) |
| [26] |
MARTIN C A, STABLER L B. Plant gas exchange and water status in urban desert landscapes[J]. J Arid Environ, 2002, 51(2): 235-254. DOI:10.1006/jare.2001.0946 ( 0) |
| [27] |
SCHOLES J D, PRESS M C, ZIPPERLEN S W. Differences in light energy utilisation and dissipation between dipterocarp rain forest tree seedlings[J]. Oecologia, 1996, 109(1): 41-48. ( 0) |
| [28] |
刘金祥, 麦嘉玲, 刘家琼. CO2浓度增强对沿阶草光合生理特性的影响[J]. 中国草地, 2004, 26(3): 14-18. ( 0) |
| [29] |
余淑文. 植物生理学和分子生物学[M]. 北京: 科学出版社, 1992: 236-240.
( 0) |
| [30] |
王会肖, 刘昌明. 作物水分利用效率内涵及研究进展[J]. 水科学进展, 2000, 11(1): 99-104. DOI:10.3321/j.issn:1001-6791.2000.01.018 ( 0) |
| [31] |
葛滢, 常杰, 陈增鸿. 青冈(Quercus glauca)净光合作用与环境因子的关系[J]. 生态学报, 1999, 19(5): 683-688. DOI:10.3321/j.issn:1000-0933.1999.05.016 ( 0) |
| [32] |
宋清海, 张一平, 郑征, 等. 热带季节雨林冠层树种绒毛番龙眼的光合生理生态特性[J]. 应用生态学报, 2006, 17(6): 961-966. DOI:10.3321/j.issn:1001-9332.2006.06.002 ( 0) |
| [33] |
戚秋慧, 盛修武, 姜恕. 内蒙羊草草原群落光合速率日变化及其与环境条件之间相互关系的初步研究[J]. 生态学报, 1983, 3(4): 333-340. ( 0) |
| [34] |
郭春燕, 李晋川, 岳建英, 等. 两种高质牧草不同生育期光合生理日变化及光响应特征[J]. 生态学报, 2013, 33(6): 1751-1761. ( 0) |