 2015, Vol. 36
2015, Vol. 36



2. 九台市水产技术推广站,吉林 九台 130599;
3. 长春通威饲料公司,吉林 长春 130404
2. Jiutai Fisheries Technology Extension Station, Jiutai 130599, China;
3. Tongwei Changchun Feed Company Limited, Changchun 130404, China
鱼类对蛋白质的代谢和利用过程包括消化、吸收、运输、分解、合成和排泄等复杂环节,受到酶、激素和基因的调控.饲料品质影响蛋白质的消化酶和肝胰脏转氨酶的活性及血液蛋白代谢产物.随着饲料蛋白水平的上升,宝石鲈Scortum bacoo胃蛋白酶和肠蛋白酶的活性增加[1].刘勇等[2]在对幼建鲤Cyprinus carpio var.jian试验得知,随饲料蛋白水平的提高试验鱼肝胰脏谷草转氨酶(AST)和谷丙转氨酶(ALT)活力升高.唐玲等[3]对镜鲤C.carpio、黄金凤等[4]对松浦镜鲤C.carpio Songpu、刘伟等[5]对奥尼罗非鱼Oreochromis niloticus♀×O.aureus♂的研究表明,随着饲料蛋白水平的升高,试验鱼血清中总蛋白(TP)、白蛋白(ALB)和球蛋白(GLB)随之而升高.本试验选取框镜鲤C.carp(Mirror carp)为研究对象, 在其饲料以植物蛋白为主要蛋白源时,添加赖氨酸和蛋氨酸2个主要的限制性氨基酸,探讨饲料必需氨基酸(EAA)平衡关联度对框镜鲤生长、消化和血清蛋白代谢指标的影响,为生产上降低框镜鲤饲料蛋白水平和饲料成本提供科学的依据.
1 材料与方法 1.1 试验饲料本试验采用鱼粉为主要蛋白源, 以鱼油、大豆油、糊精和面粉为能源、纤维素为填充物配制的半精制日粮为对照组饲料(蛋白质质量分数33%, 能量17 MJ·kg-1), 其EAA平衡关联度为0.764 9.各试验组以混合植物蛋白为蛋白源,分别添加不同水平的微囊蛋氨酸和微囊赖氨酸(均来自北京市星火元科技有限公司,质量分数各为50%),配成等氮等能(蛋白质质量分数31%, 能量17 MJ·kg-1)的不同EAA平衡关联度(0.704 8、0.722 0、0.737 0、0.759 1、0.778 0和0.800 8)的6组饲料.采用灰色关联分析法计算EAA(苏氨酸、缬氨酸、甲硫氨酸+胱氨酸、异亮氨酸、亮氨酸、苯丙氨酸+酪氨酸、赖氨酸、组氨酸、色氨酸和精氨酸)的平衡关联度,即饲料中10种EAA的模式与鱼体所需的10种EAA模式的接近程度,其中框镜鲤对10种EAA需要量以实测的框镜鲤肌肉EAA组成为依据,饲料原料的10种EAA的组成亦为实测数据[6-7].
饲料EAA平衡关联度设计原则及计算:饲料EAA为参评样本,鱼类白肌EAA为参考样本,所有参评样本组成一个灰色系统,第1步计算每种饲料和白肌的EAA分别占其总EAA的百分比(Xi和X); 第2步计算各组饲料中Xi和白肌的X的绝对差(Yi), 找出Yi的最大值(Ymax)和最小值(Ymin); 第3步在整个灰色系统中,计算每组饲料中每种EAA的平衡系数,即为(Ymin+0.5Ymax)/(Yi+0.5Ymax); 第4步计算每组饲料中10种EAA的平衡系数的平均数,并作为该组饲料EAA平衡关联度.
饲料原料经粉碎过60目筛,按配方称质量、均匀混合,挤压成直径为1.5 mm颗粒,晒干后置于-4℃冰柜中保存、备用.采用105℃恒温烘干失水法(GB /T6435—1994)测定水分,采用微量凯氏定氮法(GB /T6432—1994)测定粗蛋白质,采用550℃灼烧法(GB /T6438—1992)测定粗灰分,采用索氏抽提法(GB /T6433—1994)测定粗脂肪,弹式热量计测定能量,氨基酸自动分析仪测定氨基酸的含量.饲料配方及营养组成见表 1,饲料和肌肉中EAA含量的测定值见表 2.
|
|
表 1 试验饲料配方及其营养组成(干质量) Table 1 Formulation and nutritional composition of experimental diets(dry mass) |
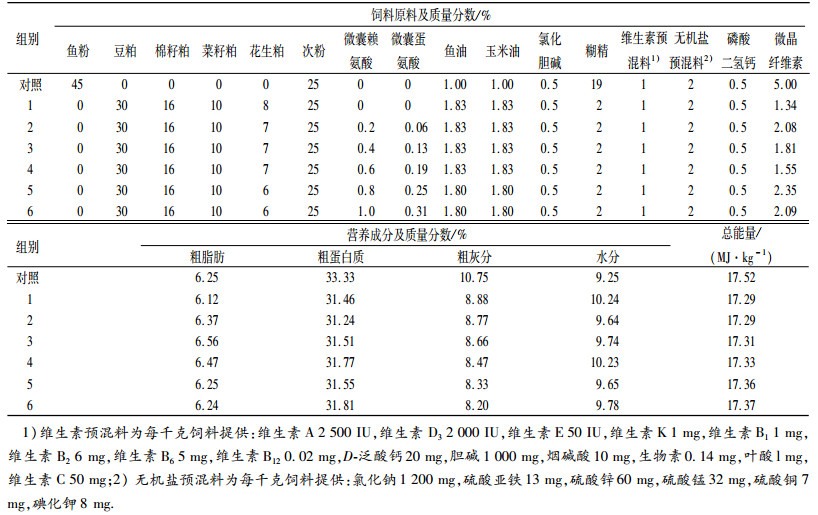
|
|
表 2 饲料和肌肉必需氨基酸质量分数(干质量) Table 2 The mass fraction of each group feed and muscle essential amino acids(dry mass) |
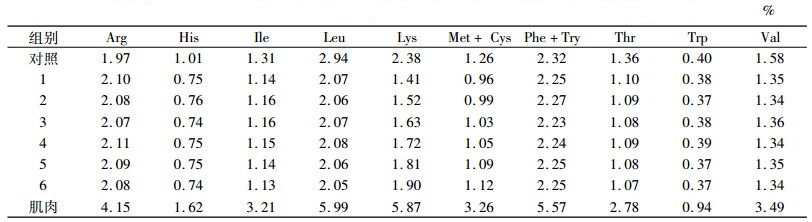
在试验前2周, 挑选规格整齐的框镜鲤鱼种1 000尾置于网箱中进行驯养,待全部摄食配合饲料后,试验开始前先停食24 h,从驯养鱼中随机取鱼放置在室内水族箱中(80 cm×60 cm×50 cm), 每种饲料3个重复, 每个重复放养30尾鱼, 每天09: 00、16: 00按2.5%~5.0%饱食投喂2次,根据鱼类的摄食进行适当调整.记录每个水族箱摄食饲料质量,试验期间水温为23~26 ℃, 溶解氧大于5.0 mg·L-1,氨氮小于0.3 mg·L-1,持续饲养8周.
1.3 测定指标 1.3.1 生长和饲料利用的指标在试验开始时测定试验鱼初始体质量,试验结束前停食24 h,测定各组试验鱼终末体质量.根据以下公式计算平均增质量率、特定生长率、饲料摄食率、饲料效率和蛋白质效率.
平均增质量率=(mt-m0)/m0×100%,
特定生长率=[(ln mt-ln m0)/t]×100%,
饲料摄食率=m料/[(m0+mt)/2×t]×100%,
饲料效率=(mt-m0)/m料×100%,
蛋白质效率=(mt-m0)/(m料×mP),
式中,m0和mt为试验开始和试验结束时鱼体的总质量(g), t为试验的时间(d), m料为摄入饲料的干质量(g), mP为饲料蛋白质的质量分数(%).
1.3.2 消化酶和代谢酶的测定饲养8周后,饥饿24 h,每组随机取15尾,生理盐水清洗肠道, 取前肠与肝胰脏, -20℃条件保存.
粗酶液制备以及蛋白酶活力测定:取前肠或肝胰脏样品, 加10倍体积的重蒸水, 4 ℃、3 000 r·min-1离心10 min,取上清液为粗酶液,4℃条件保存.采用福林-酚法测定蛋白酶活力.1个蛋白酶的比活力单位(IU或U)定义为在pH 7.0的情况下,底物酪蛋白质量浓度为20 mg·mL-1,37℃水浴10 min,每毫克酶在每分钟所产生1 μg酪氨酸的量.考马斯亮兰法测定酶液的蛋白质含量.
1.3.3 血清生化指标饲养8周后,饥饿24 h,每组取10尾鱼,用一次性注射器从尾静脉取血,4℃条件放置24 h,6 000 r·min-14℃离心10 min,取血清-20℃条件保存、待测.
血清ALT、AST、TP、ALB测定试剂盒为江苏省南京建成生物研究所产品.采用比色法测定ALT和AST的比活力,其比活力单位(IU或U)定义为1 mL血清,反应液总容量3 mL,波长340 nm,1 cm光径,25 ℃, 1 min内所生成的丙酮酸,使NADH氧化成NAD+而引起光密度每下降0.001为1个卡门氏单位,1卡门氏单位=0.482 IU·L-1.考马斯亮兰法测定酶液的蛋白质含量.
1.4 数据处理采用SPSS(18.0)软件进行单因素方差分析,用Duncan’s多重比较分析组间差异显著性程度.试验数据均以平均值±标准差表示.
2 结果与分析 2.1 饲料EAA平衡关联度对框镜鲤生长和饲料利用的影响饲料EAA平衡关联度对框镜鲤生长和饲料利用的影响见表 3.随着饲料中赖氨酸和蛋氨酸的添加,框镜鲤特定生长率有增加的趋势.即当饲料EAA平衡关联度增加到0.759 1(4组)时,框镜鲤增质量率显著高于饲料EAA平衡关联度为0.704 8、0.722 0和0.737 0的1、2和3组(P<0.05), 亦显著低于对照组(P<0.05), 随着饲料EAA平衡关联度继续增加到0.778 0(5组)和0.800 8(6组)时,5组和6组的框镜鲤特定生长率与对照组均差异不显著,但都显著高于其他各试验组(P<0.05).
|
|
表 3 饲料EAA平衡关联度对框镜鲤生长和饲料利用的影响1) Table 3 The effect of dietary EAA balance relativity on growth and feed utilization of Cyprinus carp |

随着饲料中赖氨酸和蛋氨酸的添加,饲料EAA平衡关联度对框镜鲤的摄食率没有显著影响.各试验组饲料效率随着饲料EAA平衡关联度的增加而增加,当饲料EAA平衡关联度增加到0.778 0(5组)时,饲料效率显著高于0.704 8(1组)、0.722 0(2组)和0.737 0(3组)(P<0.05), 当增加到0.800 8 (6组)时,5组和6组之间的饲料效率无显著差异,且都与对照组无显著性差异.
各试验组蛋白质效率随饲料EAA平衡关联度的增加而增加,当饲料EAA平衡关联度增加到0.759 1 (4组)时,较1、2和3组蛋白质效率显著增加(P<0.05), 当增加到0.778 0(5组)时,与饲料EAA平衡关联度0.759 1(4组)的蛋白质效率无显著差异,当增加到0.800 8(6组)时,与5组的蛋白质效率无显著性差异,但显著高于其他组(P<0.05).
2.2 饲料EAA平衡关联度对框镜鲤蛋白酶和肝胰脏转氨酶的影响饲料EAA平衡关联度对框镜鲤蛋白酶和肝胰脏转氨酶的影响见表 4.各试验组随EAA平衡关联度增加到0.737 0(3组)时,框镜鲤的肝胰脏蛋白酶比活力显著高于1和2组(P<0.05), 当EAA平衡关联度增加到0.759 1(4组)和0.778 0(5组)时,框镜鲤的肝胰脏蛋白酶比活力显著高于1~3组(P<0.05), 但与对照组差异不显著.随着饲料中EAA平衡关联度的提高,当增加到0.800 8(6组)时,框镜鲤肝胰脏的蛋白酶比活力显著高于1~4组和对照组(P<0.05).在各试验组中,饲料中EAA平衡关联度对框镜鲤肠道蛋白酶比活力没有显著性影响.各试验组肝胰脏ALT和AST酶随着饲料EAA平衡关联度的增加而增加,当饲料EAA平衡关联度增加到0.759 1(4组)时,肝胰脏ALT与对照组差异不显著.当饲料EAA平衡关联度增加到0.778 0(5组)时,肝胰脏AST显著高于1和2组(P<0.05), 且与对照组差异不显著.表明在本试验条件下,在植物蛋白源饲料中添加赖氨酸和蛋氨酸,调控饲料EAA平衡关联度为0.759 1时,框镜鲤肝胰脏蛋白酶和氨基酸转氨酶比活力达到对照组(鱼粉组)的水平,即该添加水平可促进蛋白质的消化和代谢.
|
|
表 4 饲料EAA平衡关联度对框镜鲤蛋白酶和肝胰脏转氨酶比活力的影响1) Table 4 The effect of dietary EAA balance relativity on protease and transaminase in hepatopancreas specific activity of Cyprinus carp |
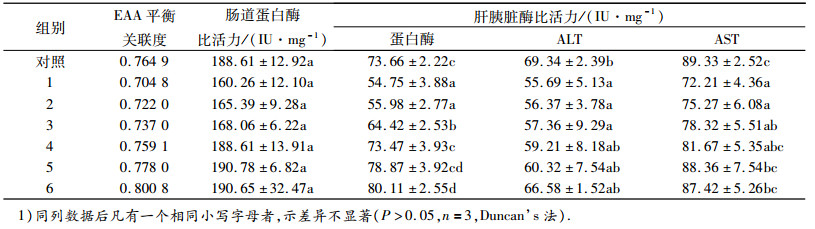
饲料EAA平衡关联度对框镜鲤血清蛋白代谢相关生化指标的影响见表 5.随着饲料中赖氨酸和蛋氨酸的添加,当饲料EAA平衡关联度增加到0.759 1 (4组)时, 血清TP与对照组差异不显著.当饲料EAA平衡关联度增加到0.778 0(5组)时,血清TP显著高于1~3组(P<0.05), 4组、5组和6组的血清TP无显著差异,且与对照组亦差异不显著.当饲料EAA平衡关联度增加到0.737 0(3组)时,血清ALB与对照组差异不显著,当饲料EAA平衡关联度增加到0.800 8(6组)时,血清ALB显著高于1和2组(P<0.05).随着饲料中赖氨酸和蛋氨酸的添加,饲料EAA平衡关联度对框镜鲤的血清ALT和AST没有显著影响.表明在本试验条件下,在植物蛋白源饲料中添加赖氨酸和蛋氨酸,调控饲料EAA平衡关联度为0.737 0~0.759 1时,框镜鲤血清蛋白代谢水平达到对照组(鱼粉组)的水平,即该添加水平可促进蛋白质的代谢.
|
|
表 5 饲料氨基酸平衡关联度对框镜鲤血清生化指标的影晌 Table 5 The effect of dietary EAA balance relativity on biochemical indicators of serum of Cyprinus carp |
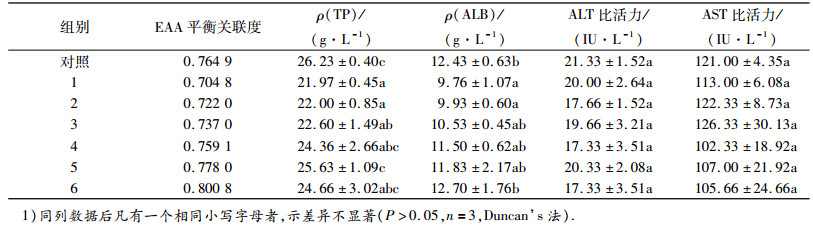
与鱼粉相比,植物蛋白源不仅EAA含量较低,还常缺乏某种或某几种EAA,蛋氨酸和赖氨酸则是其中最主要的2种限制性氨基酸[8].已有很多研究表明,饲料氨基酸的组成和平衡关联度对鱼类的生长和饲料利用有很大影响.在替代鱼粉的植物蛋白源饲料中补充添加蛋氨酸或赖氨酸对水产动物的生长和饲料利用有明显促进作用.如周长海等[9]研究表明添加合成蛋氨酸可以补充豆粕替代5%鱼粉饲料中缺乏的蛋氨酸,显著提高幼鲤的质量增加率和蛋白质效率.刘永坚等[10]在实用饲料中添加包膜赖氨酸、罗运仙等[11]在低豆粕饲料中补充微囊赖氨酸均显著改善了草鱼的生长.众所周知,鱼类对蛋白质的需求实质就是对EAA的需求,合理调整饲料中蛋氨酸和赖氨酸这2种限制性氨基酸的配比,可大大改善饲料氨基酸平衡关联度,提高蛋白源利用率. Alam等[12]研究表明在日本对虾Marsupenaeus japonicus饲料中按氨基酸最适比例同时补充包膜赖氨酸和蛋氨酸,其促生长效果优于单独补充晶体赖氨酸或蛋氨酸.于海瑞等[13]研究表明在罗非鱼Oreochromis mossambicus饲料中补充0.5%微囊赖氨酸和0.1%微囊蛋氨酸, 使得罗非鱼的蛋白质效率及饲料蛋白的表观消化率显著提高.谭芳芳等[14]在草鱼饲料中补充微囊赖氨酸和蛋氨酸,使饲料EAA平衡关联度增加,蛋白质沉积率得到提高.调控氨基酸的组成和平衡关联度不仅仅体现在这2种限制性氨基酸上,如林香信等[15]研究表明用多种外源氨基酸调节无鱼粉饲料配方氨基酸的平衡性,能够提高罗非鱼的生长性能、饲料转化率及蛋白质效率.本试验考虑到饲料中2个限制性氨基酸的含量和比例,向饲料中按比例添加蛋氨酸和赖氨酸,通过灰度关联分析,使得饲料达到不同的平衡关联度,当EAA平衡关联度为0.759 1时,框镜鲤的生长和饲料利用情况与对照组无显著差异.表明饲料中按比例添加2种主要的限制性氨基酸调控饲料EAA达到适当的平衡关联度,可提高框镜鲤生长和对饲料的利用率, 进而达到降低饲料蛋白质的水平,降低饲料成本的目的.
3.2 饲料EAA平衡关联度对框镜鲤蛋白质代谢的影响水生动物在对蛋白质的消化过程中,蛋白酶起了重要的作用,同样其血液蛋白代谢产物和氨基酸代谢酶亦反应蛋白质代谢状况和饲料对其的影响.王吉桥等[16]研究表明在适宜范围内随饲料中包膜赖氨酸添加量的增加,仿刺参Apostichopus japonicus幼参消化道中蛋白酶活力随之增加.冯琳等[17]、帅柯等[18]在对幼建鲤添加不同梯度水平蛋氨酸的试验中,得出与之相似的结论.本试验随赖氨酸与蛋氨酸在饲料中添加量的提高,EAA平衡关联度的不断提高,框镜鲤肠道蛋白酶比活力虽有增加的趋势,但差异不显著,差异的原因可能是因为添加氨基酸的种类和数量等综合因素造成的.
正常情况下,血清中转氨酶活性值较小,当肝细胞受到损伤时,细胞膜的通透性加大,大量ALT和AST渗入血液,肝的ALT和AST活性明显下降,而血液中ALT和AST活性明显升高,所以血清ALT和AST水平被认为是指示肝脏损伤的敏感指标[19].有研究认为,饲料的EAA平衡关联度影响鱼类血清和肝脏的ALT和AST活性.如叶元土等[20]试验发现,饲料中大量使用菜籽后,草鱼血清中转氨酶活性大幅升高,对其肝脏造成损伤.严全根等[21]研究发现许氏平鲉Sebastods schlegelii幼鱼肝脏AST、ALT活性随着饲料中赖氨酸水平的增加先下降,然后上升,表明当赖氨酸含量不足或过量时,氨基酸模式不平衡,部分氨基酸可能分解并以能量的形式消耗,而供应机体合成蛋白和其他生理需要的氨基酸量不足,因而导致生长和饲料利用下降.汪益峰等[22]研究发现,氨基酸平衡日粮组的异育银鲫Carassius auratus gibelio肝胰脏转氨酶浓度高于不平衡日粮组,说明氨基酸平衡提高了蛋白质的合成,从而促进异育银鲫的生长.本试验各试验组框镜鲤血清ALT和AST差异不显著,与对照组亦无显著性差异,表明饲料蛋白质量的差异和EAA平衡关联度没有影响肝胰脏的功能.而各试验组肝胰脏ALT和AST随饲料EAA平衡关联度的增加而增加,表明随着饲料蛋白质的提高促进氨基酸的代谢,从而提高蛋白质的利用率,促进蛋白质合成.
血清中TP、ALB与GLB含量可直观体现机体对蛋白质吸收和代谢情况.唐玲等[3]在对镜鲤、黄金凤等[4]在对松浦镜鲤试验得知,随着饲料蛋白含量的升高,血清中TP、ALB和GLB的浓度也升高.同样,刘伟等[5]用不同蛋白水平的饲料饲喂奥尼罗非鱼,结果随饲料蛋白水平的增加,血清ALB、GLB和TP亦随之升高,加强了蛋白质的合成与运转,从而在血清蛋白中出现了较多的蛋白质组分.本试验随饲料EAA平衡关联度的增大,血清ALB显著增加,血清TP含量明显提高,说明随EAA平衡关联度的提高,蛋白质的合成也随之提高.
在本试验条件下,在饲料中按比例添加赖氨酸和蛋氨酸调节EAA的平衡关联度,当EAA平衡关联度为0.759 1时,可促进框镜鲤的生长和饲料利用,促进蛋白质的消化和代谢,因此可适当降低饲料的蛋白含量,降低饲料成本.
| [1] |
邵庆均, 苏小凤, 许梓荣. 饲料蛋白水平对宝石鲈增重和胃肠道消化酶活性影响[J]. 浙江大学学报:农业与生命科学版, 2004, 30(5): 553-556. (  0) 0) |
| [2] |
刘勇, 周小秋, 刘扬, 等. 蛋白质对幼建鲤生长性能、消化功能和蛋白质代谢的影响[C]//刘建新. 中国畜牧兽医学会动物营养学分会第十次学术研讨会论文集. 北京: 中国农业科学技术出版社, 2008: 171.
( 0) |
| [3] |
唐玲, 徐奇友, 王常安, 等. 不同水温和饲料蛋白质水平对镜鲤血清生化指标的影响[J]. 大连海洋大学学报, 2011, 26(1): 41-46. DOI:10.3969/j.issn.1000-9957.2011.01.008 ( 0) |
| [4] |
黄金凤, 徐奇友, 王常安, 等. 温度和饲料蛋白质水平对松浦镜鲤幼鱼血清生化指标的影响[J]. 大连海洋大学学报, 2013, 2(2): 184-192. ( 0) |
| [5] |
刘伟, 文华, 蒋明, 等. 2种蛋白水平的饲料循环投喂对奥尼罗非鱼幼鱼生长、体成分和血清生化指标的影响[J]. 华南农业大学学报, 2013, 34(3): 405-412. ( 0) |
| [6] |
杨元昊, 刑西谋. 鱼用饲料中必需氨基酸平衡效果的微机分析方法研究[J]. 淡水渔业, 1999, 29(6): 6-12. ( 0) |
| [7] |
江龙建, 姜勋半, 周光宏. 应用灰色关联分析评价饲料蛋白质氨基酸平衡性[J]. 四川动物和兽医科学, 2001, 28(2): 120-124. ( 0) |
| [8] |
MURTHY H S, VARGHESE T J. Total sulphur amino acid requirement of the Indian major carp, Labeo rohita (Hamilton)[J]. Aquacult Nutri, 1998, 4(1): 61-65. DOI:10.1046/j.1365-2095.1998.00045.x ( 0) |
| [9] |
周长海, 贾友刚, 齐淑艳, 等. 低鱼粉日粮添加蛋氨酸对幼鲤生长的影响[J]. 水产科学, 2011, 30(4): 206-209. DOI:10.3969/j.issn.1003-1111.2011.04.005 ( 0) |
| [10] |
刘永坚, 田丽霞, 刘栋辉, 等. 实用饲料补充结晶或包膜赖氨酸对草鱼生长、血清游离氨基酸和肌肉蛋白质合成率的影响[J]. 水产学报, 2002, 26(3): 253-259. ( 0) |
| [11] |
罗运仙, 谢骏, 吕利群, 等. 饲料中补充晶体或微囊赖氨酸对草鱼生长和血浆总游离氨基酸的影响[J]. 水产学报, 2010, 34(3): 467-473. ( 0) |
| [12] |
ALAM M S, TESHIMA S, KOSHIO S, et al. Supplemental effects of coated methionine and/or lysine to soy protein isolate diet for junvenile kuruma shrimp, Marsupenaeus japonicus[J]. Aquaculture, 2005, 248(1/2/3/4): 13-19. ( 0) |
| [13] |
于海瑞, 张琴, 姜燕, 等. 饲料中补充赖氨酸和蛋氨酸对罗非鱼生长和消化率的影响[J]. 渔业科学进展, 2012, 33(4): 114-120. ( 0) |
| [14] |
谭芳芳, 叶元土, 肖顺应, 等. 补充微囊赖氨酸和蛋氨酸对草鱼生长性能的影响[J]. 动物营养学报, 2010, 22(3): 804-810. DOI:10.3969/j.issn.1006-267x.2010.03.042 ( 0) |
| [15] |
林香信, 陆鹏, 颜孙安, 等. 无鱼粉蛋白饲料氨基酸平衡对罗非鱼生长的影响[J]. 农学学报, 2012, 2(5): 62-66. ( 0) |
| [16] |
王吉桥, 蒋湘辉, 姜玉声, 等. 在饲料中添加包膜赖氨酸对仿刺参幼参生长、消化和体成分的影响[J]. 水产科学, 2009, 28(5): 241-245. DOI:10.3969/j.issn.1003-1111.2009.05.001 ( 0) |
| [17] |
冯琳, 周小秋, 王光花, 等. 蛋氨酸对幼建鲤肠道生长和肠道酶活的影响[C]//刘建新. 中国畜牧兽医学会动物营养学分会第十次学术研讨会论文集. 北京: 中国农业科学技术出版社, 2008: 426.
( 0) |
| [18] |
帅柯, 周小秋. 蛋氨酸对幼建鲤消化功能及免疫功能的影响[C]//冯定远. 动物营养与饲料研究: 第五届全国饲料营养学术研讨会论文集. 北京: 中国农业科学技术出版社, 2006: 169.
( 0) |
| [19] |
惠天朝, 施明华, 朱荫媚, 等. 硒对罗非鱼慢性镉中毒肝抗氧化酶及转氨酶的影响[J]. 中国兽医学报, 2000, 20(3): 264-266. DOI:10.3969/j.issn.1005-4545.2000.03.018 ( 0) |
| [20] |
叶元土. 饲料必需氨基酸的平衡效果对草鱼生长的影响[J]. 饲料工业, 1999, 20(3): 39-41. ( 0) |
| [21] |
严全根, 解绶启, 雷武, 等. 许氏平幼鱼的赖氨酸需求量[J]. 水生生物学报, 2006, 30(4): 459-465. DOI:10.3321/j.issn:1000-3207.2006.04.015 ( 0) |
| [22] |
汪益峰, 周维仁, 章世元, 等. 氨基酸平衡和外源酶对异育银鲫的生长、氮代谢及血液生化指标的影响[J]. 江苏农业学报, 2010, 26(1): 130-135. DOI:10.3969/j.issn.1000-4440.2010.01.025 ( 0) |