 2015, Vol. 36
2015, Vol. 36



2. Center for Integrated Pest Management,North Carolina State University,1730 Varsity Dr. Suite 110,Raleigh,NC 27606,USA
2. Center for Integrated Pest Management, North Carolina State University, 1730 Varsity Dr. Suite 110, Raleigh, NC 27606, USA
柚喀木虱Cacopsylla citrisuga Yang&Li属半翅目Hemiptera胸喙亚目Sternrrhyncha木虱科Psyllidae喀木虱属Cacopsylla[1].该虫在云南德宏主要为害柠檬Citrus limon (L.) Burm.柚子Citrus maxima等芸香科柑橘属果树和枳壳,密度较高时会造成果树叶片脱落和枝干干枯等症状,严重影响果树长势[2]。杨集昆和李法圣[3] 1984在云南省保山地区腾冲县和德宏州瑞丽县记录过该虫,鲜见该虫在其他地区的报道.柚喀木虱是柑橘黄龙病(Huanglongbing,HLB)亚洲种的携带者[4],主要分布于海拔1 000~1 500 m [2],而HLB的主要为害种亚洲柑橘木虱Diaphorina citri Kuwayama分布的海拔相对较低[5],鲜见它们同时危害同一片果园的报道.
分子鉴定技术检测不受组织特异性、发育阶段等影响,并且具有数量大、多态性高、遗传稳定等优势,因此在系统发育、种群遗传分化以及谱系生物地理学研究等方面应用越来越广泛[6].例如利用RAPD技术对近缘种及隐蔽种进行鉴定[7]; 基于线粒体COI基因片段的生物条形码(Barcoding)技术被广泛应用于近缘种的鉴定和种内多样性的分析[8-10].
2013年在云南石屏县的1株椪柑上发现柚喀木虱与柑橘木虱同时为害.本文利用RAPD技术对这2种木虱若虫进行鉴定,并利用线粒体细胞色素氧化酶COI基因5'端生物条形码区域片段对云南2个地区的柚喀木虱种群进行描述,旨在从分子生物学的角度对柚喀木虱进行鉴定,并分析2个地点柚喀木虱种群的遗传变异和多样性.
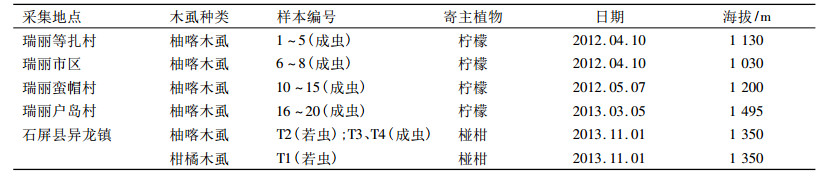
1 材料方法 1.1 供试虫源及标本处理所有虫样均采自云南瑞丽市和石屏县,采集相关信息如表 1所示.采集到的成虫和若虫均用φ为75%的乙醇溶液浸泡并编号单头保存于4℃.为了保存虫体的标本,单头成虫仅用3条足进行DNA提取,其余部分对应编号保存; 若虫根据形态鉴定并拍照,确认是柚喀木虱或柑橘木虱后,单头提取DNA.凭证标本和提取的DNA均存放于华南农业大学昆虫生态研究室.柚喀木虱形态学根据杨集昆和李法圣[3]的方法进行鉴定.
|
|
表 1 柚喀木虱和柑橘木虱样本采集信息 Table 1 Information about the samples of Cacopsylla citrisuga and Diaphorina citri |
使用天根血液/细胞/组织基因组DNA提取试剂盒(天根生化科技有限公司,北京)对单头木虱的总DNA进行提取,步骤参照试剂盒说明书.
采用引物OPA09[10]对柑橘木虱和柚喀木虱若虫(T1、T2)样本进行RAPD扩增,并以华南农业大学昆虫学实验室原有保留的柑橘木虱若虫(D1)和柚喀木虱若虫(D2)标本作为对照. RAPD扩增采用优化条件[10],扩增产物电泳后得基因组扩增图谱,进行后续数据分析.
线粒体COI片段的扩增,采用引物对CPF4和CPR4[11].聚合酶链式反应(PCR)扩增的条件如下: 94℃ 3 min的预变性; 94℃变性45 s,45.5℃退火1 min,72℃延伸1 min,35个循环; 最后72℃延伸10 min.扩增产物4℃保存.扩增产物交由华大基因科技公司纯化后,采用3730型DNA测序仪进行测序.
1.3 RAPD-PCR数据分析电泳得到的RAPD基因组扩增图谱中,每一条带代表引物与模板某一区域在一定程度上的配对.参照张丽娜[10]的方法对相同电泳条件下扩增出的2种木虱各自的特征条带进行统计,模糊不清的予以排除.
1.4 COI序列的校对、比对及系统发育分析测序得到的COI序列,在MEGA 5.1软件进行手工检查和序列的拼接,确定无移码突变和终止突变,然后进行序列比对.
通过BLAST分析在Genbank中找到的柚喀木虱COI基因的近缘序列,种属及对应的Genbank登录号为C. fraudatrix1 (JX987969)、C. fraudatrix2 (JX987968)、C. fraudatrix3 (JX987970)、C. ledi1 (JX987971)、C. ledi2 (JX987972)、C. ledi3 (JX987973)、C. myrtilli1 (KF494328)、C. myrtilli2 (KF494329)、C. myrtilli3 (KF494327)、C. myrtilli4 (KF494330)、C. myrtillii5 (KF494326)、C. myrtilli6 (KF494331)、C. myrtilli7 (KF494332)、C. pyricola1 (JF327693)、C. pyricola2 (JF327670)、C. pyricola3 (JF327671)、C. pyricola4 (JF327705)、C. pyricola5 (JF327706)、Baeopelma foersterl1 (JX987967)、Baeopelma foersterl2 (JX987966)和Bactericera cockerelli (KC008074).利用MEGA 5.1进行系统发育分析,构建邻接树,并且计算它们的种间及种内遗传距离.核苷酸替代模型选用Kimura双参数模型(K2P).对于每个节点的置信度,通过自展检验(Bootstrap)重复1 000次抽样计算.把下载的近缘序列设为外群.
1.5 柚喀木虱线粒体COI单倍型的鉴定采用Dnasp ver. 5.0软件进行单倍型的鉴定,并对变异位点数量、单倍型数量、单倍型多样度(h)、核酸多样度(π)和不同单倍型间核酸差异的均值进行统计.
2 结果与分析 2.1 RAPD分析鉴定结果若虫RAPD扩增图谱见图 1,T1和D1(对照)道发现了亚洲柑橘木虱若虫的特有条带A1和A3,T2和D2中发现了柚喀木虱所特有的条带A2.

|
图 1 引物OPA09对柑橘木虱和柚喀木虱若虫的RAPD-PCR扩增图谱 Figure 1 The RAPD-PCR amplification patterns with the primer of OPA09 on the nymphs of Diaphorina citri and Cacopsylla citrisuga M: DS2000 Marker; T1、D1(对照)为柑橘木虱,T2、D2(对照)为柚喀木虱. |
经RAPD分析鉴定,云南石屏县异龙镇的椪柑上,同枝同梢为害的2种木虱与形态学鉴定的结果一致,分别为柑橘木虱和柚喀木虱(图 2).

|
图 2 柑橘木虱(T1)和柚喀木虱(T2)若虫 Figure 2 Nymphs of Diaphorina citri (a) and Cacopsylla citrisuga (b) |
本试验获得的22条柚喀木虱线粒体COI序列,经比对后长度为622 bp.仅有1个突变核苷酸位点发生颠换(A→T),位于序列75 bp处.所有序列中只有2条序列发生了该突变,A→T颠换位于第3密码子,造成氨基酸由苯丙氨酸变为亮氨酸.碱基T、A、C、G的平均含量分别为36.5%、28.9%、20.4%、14.1%,存在明显的AT偏向性.序列被Genbank收录,登陆号为KJ850285~850306.
2.3 遗传距离由于仅从Genbank下载到1条B. cockerelli的COI序列,因此无法计算其种内遗传距离. C. citrisuga、C. myrtilli与C. pyricola的种内遗传距离分别是0~0.17%、0~0.68%和0~0.17%. C. fraudatrix、C. ledi与B. foersteri的种内遗传距离皆为0.而本研究所涉及的这7种木虱的种间遗传距离范围为3%~24%.最大种内遗传距离明显小于最小种间遗传距离(图 3),说明线粒体COI基因5'端片段能很好地用于描述C. citrisuga.

|
图 3 7种木虱种内及种间遗传距离分布图 Figure 3 Distributions of the intraspecific and interspecific genetic distances of 7 psyllid species |
由于在Genbank上获得的外群COI序列扩增长短不一,加入本研究的22条柚喀木虱进行比对之后,最终用于系统发育分析长度为593 bp. 图 4是基于COI序列构建的7种木虱样本的邻接树,其中包括22条柚喀木虱序列和21条外群序列.邻接树上每个种都分别聚集成1个分支,共形成7个明显的分支,分别为C. citrisuga、C. fraudatrix、C. ledi、C. myrtilli、C. pyricola、Be. foersterl、Bc. cockerelli,且具有较高的自展值.柚喀木虱有2个分支,其中1个包括T2和T4,另1个包括其余样本.

|
图 4 基于COI序列构建的邻接树 Figure 4 The neighbor-joining tree inferred from the COI sequences 节点前的数值为自展值(仅显示≥50的值) |
共鉴定得到2种柚喀木虱线粒体COI单倍型C1和C2,除了T2和T4为单倍型C1,其余均为单倍型C2.本研究所有柚喀木虱样本的单倍型多样度(h)为0.173 ± 0.101,核酸多样度(π)为0.000 280 ± 0.000 160.其中瑞丽的各柚喀木虱种群中仅存在唯一的单倍型C1,因此单倍型多样度和核酸多样度均为0;而石屏县的柚喀木虱种群有单倍型C1和C2,单倍型多样度为0.667 ± 0.314,核酸多样度为0.001 070 ± 0.000 510.
3 讨论与结论柚喀木虱遗传分析结果显示,COI基因最大的分歧度仅为0.16%,2种单倍型之间只有1个核苷酸的差异.与Kang等[9]研究韩国的梨喀木虱COI的最大序列分歧度0.15%很接近.而其他类似的研究也有利用COI片段对一个地区同种昆虫的不同种群进行分析[12-14],一般约为1%,对比之下,柚喀木虱的最大序列分歧度显得较低.甚至有研究表明,在美洲,柑橘木虱同区域COI片段的最大序列分歧度为6%[15].与黄龙病的媒介昆虫柑橘木虱相比,柚喀木虱的最大序列分歧显得非常低,说明其种群分化水平较低.
物种的基因多样性是由多种因素控制的.一个规模巨大而且稳定的种群是维持基因多样性的重要来源[16].柚喀木虱是云南地区为害柠檬等柑橘属植物的主要害虫,其数量较多[2],COI序列出现如此低的分歧度是相当意外的.本研究采自瑞丽不同地区的柠檬植株上的柚喀木虱种群,未发现任何COI序列的变异,说明瑞丽的柚喀木虱在柠檬这种寄主上未发生区域性的分化.因此如此单一的种群很可能是其他的一些因素导致的.有研究者曾提出种群经常出现灭绝和重新繁殖会导致遗传多样性降低[17].应用这个理论,柚喀木虱种群可能经历了经常性的灭绝和繁殖的事件.满足这样的条件,有一种可能是化学杀虫剂的使用.但化学防治的杀灭能力有限[18],似乎不能让种群经常性灭绝后重新繁殖,因此这可能不是造成COI基因匀质性的原因.另一个较为合理的解释柚喀木虱可能是近期从其他地区扩散入侵的.自杨集昆和李法圣[3] 1984年首次在云南省保山地区腾冲县和德宏州瑞丽县记录过该虫,至今鲜见在其他地区发生的报道.而一些研究者认为柚喀木虱与蒋元晖等[18] 1981年在四川宁南、德昌和云南发现的高海拔发生的合叶木虱Psylla sp.为害特征极为相似,估计为同一个种[2, 19].云南周围除了其北部与四川、西藏和贵州等高原地区相连外,还在西南方向与越南、缅甸等高海拔地区相接壤.柚喀木虱是否是从这些地区和国家入侵云南有待进一步调查和研究.
本研究于云南石屏县异龙镇海拔约1 350 m地区的椪柑嫩梢上,发现柚喀木虱和柑橘木虱若虫同时为害的现象.椪柑植株上分化出2种柚喀木虱的单倍型C1和C2,其中1头为C1,2头为C2,是否是柚喀木虱扩散后,由于寄主植物的改变而导致这种分化的形成,有待进一步研究.
石屏位于瑞丽的东北方向,且相距不远,而种群遗传多样性明显高于瑞丽.若柚喀木虱从石屏扩散到瑞丽,那么很难解释为什么瑞丽的单倍型如此单一,调查的4个种群中均不存在单倍型C2.合理的解释是:柚喀木虱是由瑞丽扩散至石屏后,在寄主椪柑上繁殖后分化出新的单倍型.这似乎更支持柚喀木虱是从西南方向入侵中国的推论.
研究者曾经提出“Barcoding Gap”的概念,也就是指序列在种间和种内存在一段差距[20-21].在本研究中,柚喀木虱与木虱科的另外6个种(其中4个种属于喀木虱属)的COI基因种间距离和种内距离落在2个区间,种内遗传距离明显小于种间距离.另外,根据COI序列构建的系统发育分析结果显示,7种木虱都各自聚成一支.因此,遗传距离分析和系统发育分析说明COI基因5'端区域作为条形码序列,具备了将柚喀木虱与其他木虱甚至同属木虱区分的能力.
| [1] |
李法圣. 中国木虱科[M]. 北京: 科学出版社, 2011.
(  0) 0) |
| [2] |
郭俊, 岑伊静, 王自然, 等. 柚喀木虱的形态、生物学特性及发生规律研究[J]. 华南农业大学学报, 2012, 33(4): 475-479. ( 0) |
| [3] |
杨集昆, 李法圣. 云南木虱科九新种及一新属[J]. 昆虫分类学报, 1984(4): 251-266. ( 0) |
| [4] |
CEN Y J, ZHANG L N, XIA Y L, et al. Detection of 'Candidatus Liberbacter asiaticus'in Cacopsylla(Psylla) citrisuga(Hemiptera: Psyllidae)[J]. Fla Entomol, 2012, 95(2): 303-309. ( 0) |
| [5] |
陈学新. 昆虫分子系统学和进化[M]//程家安, 唐振华. 昆虫分子科学. 北京: 科学出版社, 2000: 3-24.
( 0) |
| [6] |
刘春林, 游兰韶, 罗庆怀, 等. 中国棉田和稻田茧蜂近缘种的RAPD分子标记研究[J]. 动物分类学报, 2003, 8(4): 568-572. DOI:10.3969/j.issn.1000-0739.2003.04.003 ( 0) |
| [7] |
HEBERT P D N, RATNASINGHAM S, DEWAARD J R. Barcoding animal life: Cytochrome coxidase subunit 1 divergences among closely related species[J]. P Roy Soc B-Bio Sci, 2003, 270(S): 96-99. ( 0) |
| [8] |
BLAXTER M, MANN J, CHAPMAN T, et al. Defining operational taxonomic units using DNA barcode data[J]. P Roy Soc: B Bio Sci, 2005, 360(1462): 1935-1943. ( 0) |
| [9] |
KANG A R, BAEK J Y, LEE S H, et al. Geographic homogeneity and high gene flow of the pear psylla, Cacopsylla pyricola(Hemiptera: Psyllidae), detected by mitochondrial COI gene and nuclear ribosomal internal transcribed spacer 2[J]. Anim Cells Syst, 2012, 16(2): 145-153. DOI:10.1080/19768354.2011.607511 ( 0) |
| [10] |
张丽娜. 黄龙病亚洲种的两种媒介昆虫及其遗传多样性[D]. 广州: 华南农业大学, 2012. http://www.wanfangdata.com.cn/details/detail.do?_type=degree&id=Y2245250
( 0) |
| [11] |
BAE J S, KIM I, KIM S R, et al. Mitochondrial DNA sequence variation of the mushroom pest flies, Lycoelia mali (Diptera: Sciaridae), and Coboldia fuscipes (Diptera: Scatopsidae), in Korea[J]. Appl Entomol Zool, 2001, 36(4): 451-457. DOI:10.1303/aez.2001.451 ( 0) |
| [12] |
KIM K Y, JANG S K, PARK D W, et al. Mitochondrial DNA sequence variation of the tiny dragonfly, Nannophya pygmaea(Odonata: Libellulidae)[J]. Int J Indust Entomol, 2007, 15(1): 47-58. ( 0) |
| [13] |
JEONG H C, KIM J A, IM H H, et al. Mitochondrial DNA sequence variation of the swallowtail butterfly, Papilio xuthus, and the cabbage butterfly, Pieris rapae[J]. Biochem Genet, 2009, 47(3/4): 165-178. ( 0) |
| [14] |
DE LEÓN J H, SÉTAMOU M, GASTAMINZA G A, et al. Two separate introductions of Asian citrus psyllid populations found in the American Continents[J]. Ann Entomol Soc Am, 2011, 104(6): 1392-1398. DOI:10.1603/AN11086 ( 0) |
| [15] |
FRANKHAM R. Relationship of genetic variation to population size in wildlife[J]. Conserv Biol, 1996, 10(6): 1500-1508. DOI:10.1046/j.1523-1739.1996.10061500.x ( 0) |
| [16] |
MARUYAMA T, KIMURA M. Genetic variability and effective population size when local extinction and recolonization of subpopulations are frequent[J]. P Natl Acad Sci USA, 1980, 77(11): 6710-6714. DOI:10.1073/pnas.77.11.6710 ( 0) |
| [17] |
UNRUH T R. Genetic structure among 18 west coast pear psylla populations: implications for the evolution of resistance[J]. Am Entomol, 1990, 36(1): 37-43. DOI:10.1093/ae/36.1.37 ( 0) |
| [18] |
蒋元晖, 李世菱, 邱柱石, 等. 一种近年发现的柑橘害虫:柑橘合叶木虱[J]. 中国柑橘, 1981(4): 30-31. ( 0) |
| [19] |
HEBERT P D N, PENTON E H, BURNS J M, et al. Ten species in one: DNA barcoding reveals cryptic species in the neotropical skipper butterfly Astraptesful gerator[J]. P Natl Acad Sci USA, 2004, 101(41): 14812-14817. DOI:10.1073/pnas.0406166101 ( 0) |
| [20] |
MEYER C P, PAULAY G. DNA barcoding: Error rates based on comprehensive sampling[J]. PLoS Biol, 2005, 3(12): 2229-2238. ( 0) |
| [21] |
MEIERR, ZHANG, ALIF. The use of mean instead of smallest interspecific distances exaggerates the size of the "barcoding gap"and leads to misidentification[J]. Systematic Biol, 2008, 57(5): 809-813. DOI:10.1080/10635150802406343 ( 0) |