 2015, Vol. 36
2015, Vol. 36



2. 华南农业大学 广东省兽医临床重大疾病综合防控重点实验室,广东 广州 510642
2. Key Laboratory of Comprehensive Prevention and Control for Severe Clinical Animal Diseases of Guangdong Province, South China Agricltural University, Guangzhou 510642, China
2006年,全新的甲型H3N2亚型犬流感病毒(Canine influenza virus,CIV)在中国广东省被分离,经过基因组序列比对分析显示为禽源性的流感病毒[1].随后的研究发现H3N2亚型犬流感病毒在犬群中有很高的发病率,该病毒在犬群中流行并可能对犬以及人类健康造成威胁[2-4].
面对流感疫情,目前预防和控制主要的方法是接种疫苗和使用抗病毒药物,但是甲型流感病毒易发生变异和重组,并且疫苗的研制相对滞后,导致流感疫苗的作用有限,尤其是在流行的早期,流感的防治必须依靠药物[5-6].金刚烷衍生物(金刚烷胺等)和神经氨酸酶抑制剂(奥司他韦等)是目前使用的预防和治疗流感的药物.金刚烷胺等是M2离子通道阻断剂,能抑制病毒在感染细胞中的复制[7];奥司他韦(达菲)等是通过抑制神经氨酸酶的活性,防止病毒从感染的细胞中释放出来[8].但是这些药物毒性大,价格高,甚至可能有严重的不良反应,广泛应用还产生了一定程度的耐药性,因此有必要开发新型的抗流感药物[9].
H3N2亚型犬流感病毒属于正黏病毒科的单股负链RNA病毒,为I型包膜病毒,进入靶细胞的过程需要膜糖蛋白血凝素(Hemagglutinin,HA)的介导[10].HA分子由HA1和HA2亚基构成.HA1与靶细胞膜上的唾液酸受体相结合使病毒附着在宿主细胞上,在病毒的侵入过程中起关键的作用,因此HA1蛋白可作为靶蛋白用来筛选流感病毒侵入细胞的抑制剂.噬菌体展示技术(Phage display)是近年来出现的一种新的分子生物学技术,它可对含有数以亿计克隆的随机肽库进行快速的高通量筛选,用于与分子识别有关的领域,已经成为抗病毒制剂筛选、抗原表位分析、多肽药物研究和分子诊断试剂开发等强有力的工具[11-13].
本研究旨在运用噬菌体随机七肽库对纯化的H3N2亚型犬流感病毒HA1蛋白进行4轮亲和筛选,使用ELISA方法和抗病毒试验验证,以获得具有HA1蛋白高亲和力并且能够抑制H3N2亚型流感病毒复制的多肽,为开发抗流感病毒药物和诊断试剂奠定基础.
1 材料与方法 1.1 试验材料流感病毒毒株H3N2亚型犬流感(CIV)A/canine/Guangzhou/01/2006和H3N2亚型禽流感(AIV)A/Duck/Guangdong/W12/2011由华南农业大学兽医学院外科实验室保存;H3N2亚型犬流感病毒HIS-HA1融合表达蛋白HA1相对分子质量约为58 000,由华南农业大学兽医学院外科实验室表达及纯化[14],能与H3N2亚型CIV阳性血清发生特异性反应;MDCK细胞由华南农业大学兽医学院外科实验室保存,培养于含体积分数为10%胎牛血清的DMEM培养液;9日龄SPF鸡胚购自梅里亚维通实验动物技术公司;噬菌体随机七肽库Ph.D-7TM为New England Biolabs公司产品;M13 HRP标记单克隆抗体为GE Healthcare公司产品;CCK8试剂盒为凯基生物技术公司产品,其他试剂均为国产分析纯.
1.2 试验方法 1.2.1 噬菌体随机七肽库筛选以纯化的H3N2亚型犬流感病毒HA1蛋白作为靶分子对随机七肽库进行筛选.将HA1蛋白稀释于NaHCO3(0.1 mol·L-1,pH 8.6)包被液中至质量浓度为10 μg·mL-1,取200 μL包被酶标板,置于湿盒中4 ℃孵育过夜,链霉亲和素(0.1 mg·mL-1,100 μL)作为对照同样包被.弃包被液,加满封阻液,4 ℃作用1 h.TBST(TBS +体积分数为0.1% Tween-20)缓冲液洗涤6次后,取10 μL噬菌体随机七肽库(2 × 1011PFU)稀释于200 μL TBST缓冲液中,室温孵育1 h.TBST洗涤6次后,用0.2 mol·L-1 Glycine-HCl(pH 2.2)洗脱结合噬菌体,并用1 mol·L-1 Tris-HCl(pH 9.1)中和.吸取1 μL洗脱液测定噬菌体滴度,余下洗脱液吸取5 μL进行扩增并测定扩增后噬菌体滴度.共进行4轮筛选,每轮包被的抗原量依次递减,分别为10、5、1、1 μg·mL-1;TBST缓冲液中Tween-20的体积分数依次递增,分别为0.1%、0.2%、0.5%、0.5%.
1.2.2 筛选产物与靶蛋白HA1结合活性的ELISA鉴定将第4轮筛选得到的噬菌体库10倍系列稀释,感染大肠埃希菌ER2738后铺于双层琼脂板中间.从少于100个蓝色噬菌斑的平板上随机挑取30个,分别进行扩增纯化.纯化后的噬菌体一部分提取核酸用于测序,另一部分滴度测定后用于ELISA检测.将HA1蛋白以1 μg·mL-1包被ELISA板,对4轮筛选的洗脱产物和挑出的30个噬菌体克隆取2 × 1011PFU进行ELISA检测,噬菌体克隆进行10-1、10-2和10-3共3个稀释度稀释,同时以野生型M13噬菌体为阴性对照,以BSA为空白对照.
1.2.3 噬菌体阳性克隆的序列测定参照噬菌体展示七肽库说明书,使用NaI法快速提取纯化后噬菌体的单链DNA,并以此为模板,PCR扩增噬菌体展示多肽的DNA片段送交华大基因测序,测序引物为5′-GTATGGGATTTTGCTAAACAAC-3′.根据DNA测序结果推导出噬菌体展示多肽的氨基酸序列.
1.2.4 多肽的合成与纯化本试验中所用的多肽均由北京中科亚光生物科技有限公司合成并纯化.
1.2.5 多肽对SPF鸡胚中流感病毒繁殖的影响取50 μL血凝单位为8的CIV和AIV分别与50 μL不同浓度的亲和多肽混合并于室温孵育1 h,将混合液注入9 ~ 11日龄鸡胚中,测定病毒滴度,PBS缓冲液作为阴性对照.
1.2.6 多肽在细胞水平对流感病毒复制的影响选取对数期生长的MDCK细胞,铺96孔板,每孔5 000个细胞,置于37 ℃、体积分数为5%的CO2细胞培养箱培养24 h.取不同浓度的多肽和100 TCID50的CIV和AIV混合,室温作用1 h后接种于MDCK细胞,37 ℃孵育1 h后用PBS缓冲液洗脱未吸附的病毒,加入含1 μg·mL-1 TPCK胰酶的DMEM细胞维持液,置培养箱中培养48 h,每孔重复3次,用CCK8法检测各孔细胞存活率.
1.2.7 细胞毒性试验96孔板中每孔接种约5 000个MDCK细胞,置于37 ℃、体积分数为5% CO2的细胞培养箱中培养24 h,以含有系列稀释度待测多肽的维持液替换培养液,继续培养48 h,光学显微镜下观察细胞形态并于每孔中加入5 mg·mL-1的CCK8溶液10 μL,继续培养4 h,测定D450 nm值.每个浓度设4孔平行,计算平均值用于评价被测多肽对细胞的毒性.
1.2.8 统计学分析所有试验至少设3个独立的重复.试验数据以平均值±标准差表示,用SigmaPlot 12.2软件进行统计分析.试验组与对照组样本平均数采用单因素方差分析.
2 结果与分析 2.1 噬菌体随机七肽库的筛选以纯化的H3N2亚型犬流感病毒HA1蛋白为作用靶分子对噬菌体随机七肽库进行4轮亲和筛选,每轮筛选前加入的噬菌体量和洗脱后的噬菌体量进行比较,4轮筛选噬菌体投入产出比逐渐增高(表 1),说明噬菌体得到了很好的富集.ELISA鉴定结果显示每轮洗脱产物与HA1蛋白的亲和力逐步提高(图 1),经过第4轮筛选后的噬菌体展示肽库产率和结合力没有很大的提高,说明富集作用已达到最高,故停止筛选.
|
|
表 1 亲和筛选噬菌体的产率 Table 1 Yield ratios of the phage by biopanning |
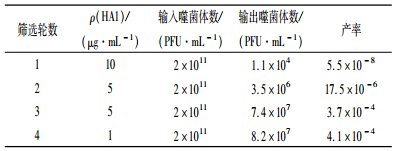

|
图 1 每轮洗脱产物多克隆噬菌体展示随机多肽与HA1蛋白的亲和力 Figure 1 Binding of polyclonal phage display peptides to protein HA1 from round 1 to 4 |
第4轮筛选后,从少于100个蓝色噬菌斑的双层琼脂板中挑选30个进行ELISA鉴定,采用BSA为空白对照,野生型M13噬菌体为阴性对照,所测D450 nm高于阴性对照3倍以上即可认定为阳性克隆. ELISA结果显示挑取的30个克隆中有17个是阳性克隆,阳性噬菌体克隆的HA1蛋白亲和力如图 2所示(部分数据未列出),野生型M13的D450 nm很小,而阳性单个噬菌体克隆随着稀释度的升高,D450 nm逐渐增大,说明6个噬菌体克隆与HA1蛋白具有较高的亲和力.

|
图 2 第4轮筛选的6个阳性噬菌体克隆不同稀释度的HA1蛋白亲和力ELISA检验结果 Figure 2 Detection of protein HA1 with monoclonal phagedisplay peptides from round 4 by ELISA |
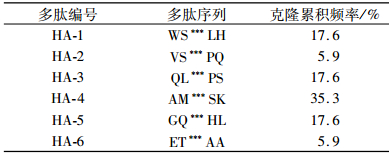
将挑选的30个蓝色噬菌斑扩增纯化后进行DNA测序,根据测出的DNA序列推导出相应的氨基酸序列,经序列比对并结合ELISA鉴定结果共获得6条不同的七肽(表 2),HA-4的克隆累积频率达到35.3%.北京中科亚光生物科技有限公司合成这6条HA1蛋白亲和肽,并使用HPLC方法纯化后用于后续试验(纯度大于98.5%).
|
|
表 2 噬菌体展示筛选得到的6条多肽序列 Table 2 Six peptide sequences selected from the phage dis- play random peptide library |
取浓度为1 000 μmol·L-1的多肽HA-1、HA-2、HA-3、HA-4、HA-5和HA-6各50 μL分别与50 μL血凝单位为8的CIV和AIV混合,室温孵育1 h后注入鸡胚尿囊腔中,进行病毒滴度测定,PBS作为阴性对照.图 3显示了6条多肽在鸡胚内具有不同程度的抗病毒活性,其中HA-3对AIV的抗病毒活性明显强于CIV;HA-4对2种流感病毒的抗病毒活性相比其他多肽最好,能够使病毒滴度下降约50%;HA-5对CIV的抗病毒活性高于AIV.

|
图 3 亲和多肽在鸡胚内抗H3N2亚型流感病毒活性验证 Figure 3 Antiviral activities of binding peptides in chicken embryos 相同柱子上,*表示与阴性对照组相比差异显著(P<0.05). |
将HA- 4稀释为5个浓度(0、1、10、100和1 000 μmol·L-1),各取5 0 μL分别与5 0 μL血凝单位为8的CIV和AIV混合,室温下孵育1 h后进行鸡胚试验,测定病毒滴度.图 4中显示了多肽HA-4对这2株病毒的繁殖都有抑制作用,在100和1 000 μmol·L-1浓度下与阴性对照(0 μmol·L-1)相比具有显著性差异,随着多肽浓度的增高流感病毒的滴度在下降,说明多肽HA-4在鸡胚水平上抑制流感病毒增殖能力逐渐增强,并且呈现剂量依赖特征.

|
图 4 多肽HA-4在鸡胚内抗H3N2亚型流感病毒活性 Figure 4 Antiviral activities of peptide HA-4 in chicken embryos 相同柱子上,*表示与阴性对照组相比差异显著(P<0.05). |
将稀释成5个浓度(0、1、10、100和1 000 μmol·L-1)的多肽HA-4与100 TCID50的CIV和AIV混匀后室温孵育1 h,接种于MDCK细胞,培养48 h后用CCK8法检测细胞存活率.结果如图 5所示,多肽HA-4对2种流感病毒都有比较好的抑制作用,随着多肽浓度的增高,MDCK细胞的存活率逐渐升高,说明多肽HA-4在细胞水平上抑制流感病毒增殖能力逐渐增强,并且呈现剂量依赖特征.

|
图 5 多肽HA-4在细胞水平抗H3N2亚型流感病毒活性 Figure 5 Antiviral activities of peptide HA-4 in vitro 相同柱子上,*表示与阴性对照组相比差异显著(P<0.05). |
为了初步探究多肽HA-4作用于病毒感染细胞的阶段,细胞接毒前将稀释成5个浓度(0、1、10、100和1 000 μmol·L-1)的多肽HA-4与100 TCID50的CIV混匀后室温孵育1 h,接种于96孔板,培养48 h后用CCK8法检测细胞存活率;100 TCID50的CIV接种于96孔板后,在细胞维持液中加入5个终浓度(0、1、10、100和1 000 μmol·L-1)的多肽HA-4,培养48 h后用CCK8法检测细胞存活率.如图 6所示,多肽HA-4接毒前处理病毒组随着多肽浓度的增高细胞的存活率逐渐增高;接毒后添加多肽组与阴性对照组(0 μmol·L-1)没有显著性差异,多肽对病毒复制的抑制作用不明显.多肽接毒前处理组比接毒后添加多肽组的细胞存活率明显提高,说明了多肽预处理组抑制流感病毒复制的作用明显,初步验证了多肽HA-4可能作用于流感病毒感染靶细胞的黏附阶段,与HA1蛋白结合阻止病毒附着靶细胞,从而发挥抗病毒作用.

|
图 6 多肽HA-4作用于病毒感染细胞阶段的验证 Figure 6 Exploration of peptide HA-4 function in the stage of virus infect cells *表示与阴性对照组相比,差异显著(P<0.05). |
为了评估多肽对细胞的毒性,通过CCK8方法测定了不同浓度下HA-4对MDCK细胞的毒性.HA-4在高达1 000 μmol·L-1时仍未表现出明显的细胞毒性(图 7).

|
图 7 多肽HA-4在MDCK细胞上的细胞毒性 Figure 7 In vitro toxicities of peptide HA-4 in MDCK cells |
阐明抗原抗体反应的生物学分子作用机制,是研究正常生理学作用和疾病防治的基础和必要的步骤.流感病毒HA蛋白在病毒吸附和进入靶细胞的过程中起非常重要的作用,近年来,发现了很多小分子多肽作用于HA蛋白而抑制流感病毒的繁殖.2006年Jones等[15]发现的1条来源于成纤维细胞生长因子-4的20个氨基酸的多肽(EB),具有广谱的抗流感病毒活性,作用机理是作用于流感病毒的HA蛋白阻止病毒进入细胞.2009年Rajik等[16]运用噬菌体随机肽库针对禽流感H9N2全病毒颗粒筛选出1条多肽,经鉴定作用于HA蛋白,对于H9N2亚型禽流感有非常显著的抗病毒活性.2013年,汪崇文等[17]以H1N1型流感病毒HA蛋白为作用靶点在噬菌体随机十二肽库中筛选出9条多肽,其中H6有明显的抗病毒活性.Jeon等[18]将HA1保守区域作为靶分子,通过SELEX程序筛选DNA寡核苷酸库,获得2个与靶分子特异结合最强的适应配体,在体外试验和动物模型中都能有效抑制流感病毒HA活性,而且能抑制不同禽流感病毒毒株的感染.这些抗流感病毒多肽的发现,为抗流感病毒药物的研究指明方向.
在本研究中,用H3N2亚型犬流感HA1蛋白作为靶分子进行噬菌体展示随机七肽库筛选,从而避免了使用全病毒颗粒筛选可能导致的NA以及M2蛋白的干扰[16].筛选出来的阳性噬菌体克隆对HA1蛋白有很高的亲和力,6条HA1亲和多肽对犬流感A/canine/Guangzhou/01/2006和禽流感A/Duck/ Guangdong/W12/2011病毒有不同程度的抑制活性,且成剂量依赖特性.这些多肽可能模拟不连续的结合位点,与HA1蛋白从不同位置形成一个重要的接触面[19].推断出多肽可以作用于HA蛋白的保守区域,从而抑制流感病毒与靶细胞受体的结合[20].竞争ELISA试验证实了展示于噬菌体上的七肽段与HA1蛋白结合而不是噬菌体的其他部分.阳性噬菌体克隆的HA1蛋白亲和力与亲和多肽的抑制病毒复制能力不呈现平行关系,试验显示HA-4具有最强抗病毒复制能力,但是噬菌体克隆的亲和力并不是最高.这可能与不同多肽结合在HA1蛋白的不同位点有关,亲和力高的位点不一定跟病毒的黏附和侵入有关[21].然而,需要进一步的研究来验证多肽具体的作用位点和机理,为研制高效的抗病毒分子药物和诊断试剂奠定基础.
| [1] |
LI S, SHI Z, JIAO P, et al. Avian-origin H3N2 canine influenza A viruses in Southern China[J]. Infect, Genet Evol, 2010, 10(8): 1286-1288. DOI:10.1016/j.meegid.2010.08.010 (  0) 0) |
| [2] |
LEE C, SONG D, KANG B, et al. A serological survey of avian origin canine H3N2 influenza virus in dogs in Korea[J]. Vet Microbiol, 2009, 137(3/4): 359-362. ( 0) |
| [3] |
粟硕, 李华涛, 陈济铛, 等. 犬流感(H3N2亚型)血清学调查[J]. 中国畜牧兽医, 2013, 40(2): 201-204. DOI:10.3969/j.issn.1671-7236.2013.02.048 ( 0) |
| [4] |
ZHANG Y, CHEN J, XIE J, et al. Serologic reports of H3N2 canine influenza virus infection in dogs in Northeast China[J]. J Vet Med Sci, 2013, 75(8): 1061-1062. DOI:10.1292/jvms.13-0022 ( 0) |
| [5] |
ELLEBDY A H, WEBBY R J. Influenza vaccines[J]. Vaccine, 2009, 27: D65-D68. DOI:10.1016/j.vaccine.2009.08.038 ( 0) |
| [6] |
HILLEMAN M R. Realities and enigmas of human viral influenza:Pathogenesis, epidemiology and control[J]. Vaccine, 2002, 20(25): 3068-3087. ( 0) |
| [7] |
WANG C, TAKEUCHI K, PINTO L H, et al. Ion channel activity of influenza A virus M2 protein:Characterization of the amantadine block[J]. J Virol, 1993, 67(9): 5585-5594. ( 0) |
| [8] |
MOSCONA A. Neuraminidase inhibitors for influenza[J]. New Engl J Med, 2005, 353(13): 1363-1373. DOI:10.1056/NEJMra050740 ( 0) |
| [9] |
DE JONG M D, THANH T T, KHANH T H, et al. Oseltamivir resistance during treatment of influenza A(H5N1)infection[J]. New Engl J Med, 2005, 353(25): 2667-2672. DOI:10.1056/NEJMoa054512 ( 0) |
| [10] |
SKEHEL J. An overview of influenza haemagglutinin and neuraminidase[J]. Biological, 2009, 37(3): 177-178. DOI:10.1016/j.biologicals.2009.02.012 ( 0) |
| [11] |
AZZAZY H M, HIGHSMITH W J. Phage display technology:Clinical applications and recent innovations[J]. Clin Biochem, 2002, 35(6): 425-445. DOI:10.1016/S0009-9120(02)00343-0 ( 0) |
| [12] |
MULLEN L M, NAIR S P, WARD J M, et al. Phage display in the study of infectious diseases[J]. Trends Microbiol, 2006, 14(3): 141-147. DOI:10.1016/j.tim.2006.01.006 ( 0) |
| [13] |
SERGEEVA A, KOLONIN M, MOLLDREM J, et al. Display technologies:Application for the discovery of drug and gene delivery agents[J]. Adv Drug Deliv Rev, 2006, 58(15): 1622-1654. DOI:10.1016/j.addr.2006.09.018 ( 0) |
| [14] |
张敦伟, 熊永忠, 远立国, 等. H3N2亚型犬流感病毒HA1基因的原核表达及抗原性分析[J]. 中国预防兽医学报, 2011, 33(2): 157-159. DOI:10.3969/j.issn.1008-0589.2011.02.19 ( 0) |
| [15] |
JONES J C, TURPIN E A, BULTMANN H, et al. Inhibition of influenza virus infection by a novel antiviral peptide that targets viral attachment to cells[J]. J Virol, 2006, 80(24): 11960-11967. DOI:10.1128/JVI.01678-06 ( 0) |
| [16] |
RAJIK M, JAHANSHIRI F, OMAR A R, et al. Identification and characterisation of a novel anti-viral peptide against avian influenza virus H9N2[J]. J Virol, 2009, 6(1): 74. DOI:10.1186/1743-422X-6-74 ( 0) |
| [17] |
汪崇文, 丁晓然, 杨静, 等. H1N1型流感病毒血凝素抑制肽的筛选研究[J]. 国际药学研究杂志, 2013, 40(1): 48-52. ( 0) |
| [18] |
JEON S H, KAYHAN B, BEN-YEDIDIA T, et al. A DNA aptamer prevents influenza infection by blocking the receptor binding region of the viral hemagglutinin[J]. J Biol Chem, 2004, 279(46): 48410-48419. DOI:10.1074/jbc.M409059200 ( 0) |
| [19] |
BAIR C L, OPPENHEIM A, TROSTEL A, et al. A phage display system designed to detect and study protein-protein interactions[J]. Mol Microbiol, 2008, 67(4): 719-728. DOI:10.1111/mmi.2008.67.issue-4 ( 0) |
| [20] |
MATSUBARA T, ONISHI A, SAITO T, et al. Sialic acid-mimic peptides as hemagglutinin inhibitors for anti-influenza therapy[J]. J Med Chem, 2010, 53(11): 4441-4449. DOI:10.1021/jm1002183 ( 0) |
| [21] |
CROSS K J, BURLEIGH L M, STEINHAUER D A. Mechanisms of cell entry by influenza virus[J]. Expert Rev Mol Med, 2001, 3(21): 1-18. ( 0) |