 2015, Vol. 36
2015, Vol. 36



当前全球的气候变化使得干旱问题日益严重[1].草坪草在生长发育过程中, 通过地下器官吸收土壤水分进行蒸腾作用和调节组织功能, 在土壤水分供应不足时, 会引起植株生理机能紊乱, 严重时造成草坪草死亡.抗旱性是草坪草抗逆性的一个重要指标[2-3].国内外研究表明, 不同种甚至不同品种的草抗旱性差异很大, 利用抗旱、低需水型草坪草建植的草坪能够节约用水达50%以上[4].因此, 开展草坪草抗旱研究, 对节约水资源、建设可持续的草坪生态系统有重要意义.在草坪草抗旱性评价方面, 很多方法已经得到应用, 比如草坪质量评价法、形态学鉴定法、表征永久萎蔫系数法、生理指标评价以及综合评价法[5].Huang等[6]用冠层温度、叶绿素含量、相对含水量和茎干旱生长量对7种暖季性草坪草在表层0 ~ 40 cm土壤干旱条件下的抗旱性进行了研究.吕静等[7]选取土壤含水量、叶片相对含水量、质膜透性、可溶性糖含量、丙二醛含量及脯氨酸含量等指标对4种暖季型草坪草的抗旱性作了排序.
结缕草Zoysia japonica作为华南地区主要的草坪草种, 具有低维护、耐践踏、耐瘠薄、抗病虫害以及耐盐碱等特性, 其不论在坪用性状还是抗逆性等方面都存在很大的遗传变异, 是一种很有发展潜力的草坪草[8-9].在抗旱性方面, 余晓华等[10]通过测定叶片相对含水量、质膜相对透性及脯氨酸含量的变化对4种结缕草的抗旱能力进行了排序.谢新春等[11]通过盆栽试验, 比较了兰引Ⅲ号、新品系ZS-1和ZSSJZ等3个结缕草品种(系)的抗旱性强弱.以上研究结果表明, 结缕草的抗旱性品种间存在很大差异.
本试验以3种结缕草和经过60Co-γ辐射诱变选育的6种新品系为材料, 在干旱胁迫条件下, 通过测定叶片的相对电导率、脯氨酸含量、可溶性总糖含量、相对含水量等生理指标和盖度、颜色等表观性状, 比较3种结缕草及其新品系的抗旱性, 为草坪草抗性育种提供科学依据.
1 材料与方法 1.1 材料试验材料共9种, 包括兰引Ⅲ号结缕草Zoysia japonica cv.Lanyin No.Ⅲ(LY)及其新品系LY-50-2 (LY-1)、LY-450-1(LY-2);ZS-1结缕草Zoysia japonica spp.(ZS)及其新品系ZS-40-2 (ZS-3)、ZS-50-2 (ZS-4);ZS-SJZ结缕草Zoysia japonica spp.(ZSS)及其新品系ZSS-50-1 (ZSS-5)、ZSS-60--2 (ZSS-6).其中, LY-1和LY-2是利用60Co-γ射线以0.14 Gy·min-1的强度分别在40和450 Gy剂量下辐射LY筛选出的新品系; ZS-3、ZS-4是利用同样方法辐射ZS筛选出的新品系; ZSS-5、ZSS-6是辐射ZSS筛选出的新品系.
1.2 试验方法盆栽试验设在华南农业大学农学院楼顶天台, 地处N 24°26′、E 118°04′, 属于亚热带海洋性季风气候, 年平均气温为21.8 ℃, 最热月(7月)多年平均气温为28.5 ℃, 最冷月(1月)多年平均气温为13.0 ℃, 年降雨量约为1 706.6 mm.试验材料于2013年7月18日种植于花盆中(花盆口径为20 cm、高为17 cm), 基质为普通细河沙与泥炭按质量比3:1的比例混合均匀, 每个品种(系)种植3盆, 共27盆, 正常养护管理.至2013年11月15日, 所有材料的盖度均达到100%, 之后统一修剪, 精心养护管理.继续培养20 d后, 于12月4日浇透水, 开始干旱胁迫处理, 在控水(一直不浇水)之后的第3、9和15天时取相同位置的叶片测定相关的生理指标, 叶片采集的时间为当天上午8点左右, 每盆取样2.0 g.试验过程中同时监测土壤含水量变化, 确定干旱胁迫程度, 第3、9和15天取样时的土壤含水量(w)分别为18.76 %、12.74 %和2.43 %.干旱胁迫处理结束后复水, 浇透水.
1.3 测定指标及方法土壤含水量:利用Field Scout TDR-100土壤水分速测仪监测土层表面至土层以下7.5 cm处含水量, 随机选取3处进行测定, 取平均数.叶片相对含水量(RWC):取不同处理的、剪碎混匀的新鲜叶片0.2 g, 除去表面灰尘, 称其鲜质量, 然后置于烘箱中, 105 ℃杀青15 min, 80 ℃烘干24 h, 称其干质量, 叶片相对含水量= [(鲜质量-干质量)/鲜质量] × 100 % [12], 重复3次.叶片相对电导率(REC)采用DDSJ-307型电导率仪(上海精密科学仪器有限公司)测定; 脯氨酸含量(Pro)采用磺基水杨酸提取茚三酮显色法测定; 可溶性糖含量(TSC)采用蒽酮法测定[13]; 重复3次.
盖度(Coverage):采用面积为10 cm × 10 cm的样方框, 随机放置, 方格内交叉点触及叶片的均算已覆盖.计算已覆盖的交叉点的数目所占百分数, 即为盖度.颜色(Color):在结缕草生长稳定期, 于2次施肥间期对颜色采用9分制评分方式进行打分.颜色墨绿为9分, 完全枯黄为1分, 6分为绿色(也为可以接受水平), 中间级别则依此尺度打分.
1.4 综合评价方法采用隶属函数值赋予权重法[14]进行抗旱性综合评价:
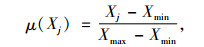
|
(1) |
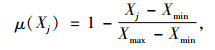
|
(2) |
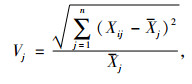
|
(3) |
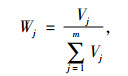
|
(4) |
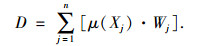
|
(5) |
式中, μ(Xj)表示第j个指标的隶属函数值, 当j指标与抗旱性正相关采用(1)式, 负相关采用(2)式; Xj表示第j个指标值; Xmin表示第j个指标最小值; Xmax表示第j个指标最大值, Xj表示第j个指标平均值; Xij表示i材料j性状的隶属函数值; Vj表示第j个指标标准差系数; Wj表示第j个指标权重; D表示各材料的综合评价值.
1.5 数据处理采用Microsoft Excel 2010进行图表分析, 采用SPSS 17.0软件进行方差分析, 采用Duncan’s法在P<0.05水平下进行多重比较.
2 结果与分析 2.1 兰引Ⅲ号结缕草及其新品系抗旱性比较由图 1A可以看出, 整个干旱胁迫时期, LY结缕草及其新品系的相对含水量(RWC)随着胁迫时间的延长呈逐渐降低的趋势, 在胁迫的第15天, LY-2的叶片RWC显著高于对照LY.LY及2个新品系随着干旱胁迫加剧叶片相对电导率(REC)逐渐增大, 但三者之间均没有显著差异(1B).在胁迫处理的第9天, LY-2的脯氨酸(Pro)质量分数显著高于LY及LY-1, 至干旱处理第15天LY的Pro质量分数达到1 638.26 μg·g-1, 显著高于LY-2和LY-1(图 1C).在干旱胁迫第9天时, LY-1的可溶性总糖(TSC)质量分数显著低于LY及LY-2, 其余阶段各品种(系)之间差异不显著(图 1D).

|
图 1 干旱胁迫对兰引Ⅲ号结缕草及新品系生理指标的影响 Figure 1 Effects of drought stress on physiological characteristics of Zoysia japonica cv.Lanyin No.Ⅲ and its new strains 各图中相同胁迫时间柱子上凡是有一个相同小写字母者, 表示差异不显著(Duncan’s法, P>0.05). |
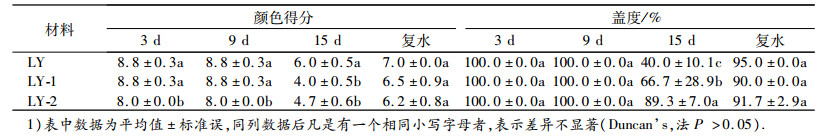
由表 1可知, 在干旱胁迫的第15天, LY结缕草及其新品系的叶片由于缺水颜色变浅, 颜色分值呈现LY>LY-2>LY-1;复水后, LY结缕草及其新品系的颜色都有所恢复, 整体呈现LY>LY-1>LY-2.LY结缕草及其新品系的盖度随着胁迫的加剧而逐渐降低, 在胁迫第15天时, 盖度呈现LY-2>LY-1>LY; 复水后, LY结缕草及其新品系恢复较快, 呈现LY>LY-2>LY-1, 但三者之间无显著差异.
|
|
表 1 干旱胁迫下兰引Ⅲ号结缕草及新品系颜色得分和盖度的变化1) Table 1 Changes of color score and coverage of Zoysia japonica Lanyin No.Ⅲ and its new strains under drought stress |
由图 2A可以看出, 整个时期ZS结缕草及其新品系RWC随着胁迫程度的增加而呈逐渐降低的趋势.在胁迫的第15天, ZS-3和ZS-4的叶片RWC显著高于ZS, 且整体呈现ZS-3>ZS-4>ZS.ZS和ZS-4随着干旱胁迫的加剧其叶片REC逐渐增大, 而ZS-3的叶片REC处于比较稳定的状态, 在干旱胁迫处理的第15 d, ZS和ZS-4叶片的REC显著高于ZS-3(图 2B).在胁迫处理的第3 d, ZS-3的Pro质量分数显著低于ZS和ZS-4, 而在胁迫的第15天, ZS-4的Pro质量分数显著高于ZS和ZS-3, 达到1 922.04 μg·g-1 (图 2C).在整个干旱胁迫时期, ZS的TSC质量分数始终高于ZS-3和ZS-4, 且在处理的第3、15天, 达到了显著水平(图 2D).

|
图 2 干旱胁迫对ZS-1号结缕草及新品系生理指标的影响 Figure 2 Effects of drought stress on physiological characteristics of Zoysia japonica 'ZS-1' and its new strains 各图中相同胁迫时间柱子上凡是有一个相同小写字母者, 表示差异不显著(Duncan’s法, P>0.05). |
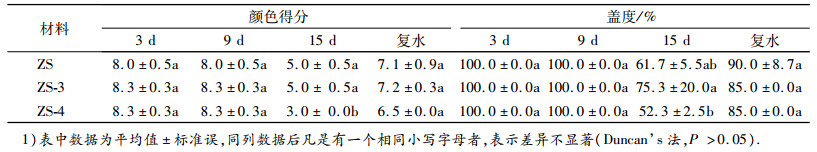
由表 2可知, 在干旱胁迫的第15天, ZS结缕草及其新品系的叶片由于缺水颜色变浅, 颜色分值呈现ZS≈ZS-3>ZS-4, 复水后, ZS结缕草及其新品系的颜色都有所恢复, 整体呈现ZS-3>ZS>ZS-4, 除第9天外, 三者之间无显著差异.ZS结缕草及其新品系的盖度随着胁迫天数的增加而逐渐降低, 在胁迫第15天时, 盖度呈现ZS-3>ZS>ZS-4, 复水后, ZS结缕草及其新品系的盖度都有所增加, 呈现ZS>ZS-3 = ZS-4, 除第9天外, 三者之间无显著差异.
|
|
表 2 干旱胁迫下ZS-1结缕草及新品系颜色得分和盖度的变化1) Table 2 Changes of color score and coverage of Zoysia japonica 'ZS-1' and its new strains under drought stress |
从图 3A可以看出, 整个干旱胁迫时期, ZSS结缕草及其新品系RWC随着胁迫程度的增加而逐渐降低.在胁迫的第15天, ZSS-5和ZSS-6的叶片RWC显著高于ZSS, 且整体呈现ZSS-6>ZSS-5>ZSS.随着干旱胁迫的加剧ZSS和ZSS-6叶片REC逐渐增大, 而ZSS-5的叶片REC则呈先升高后降低的状态, 在处理的第15天, 新品系ZSS-5的叶片REC显著低于ZSS和ZSS-6(图 3B).ZSS在干旱胁迫处理的第15天, Pro质量分数急剧增加, 达到2 326.32 μg·g-1, 显著高于ZSS-5和ZSS-6, 整个胁迫期间ZSS-5的Pro质量分数一直处于比较稳定的状态(图 3C).ZSS及其2个新品系的TSC质量分数在整个干旱胁迫期间先下降后上升, 在处理的第3天, ZSS-5的TSC质量分数显著高于ZSS-6和ZSS, 其余时期没有显著差异(图 3D).

|
图 3 干旱胁迫对ZSS-SJZ结缕草及新品系生理指标的影响 Figure 3 Effect of drought stress on physiological characteristics of Zoysia japonica 'ZSS' and its new strains 各图中相同胁迫时间柱子上凡是有一个相同小写字母者, 表示差异不显著(Duncan’s法, P>0.05). |
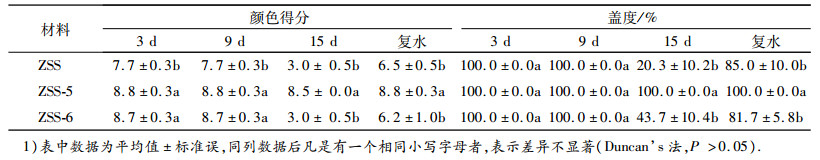
由表 3可知, 在干旱胁迫的第15天, ZSS结缕草及其新品系的叶片由于缺水颜色变浅, 颜色分值呈现ZSS-5>ZSS≈ZSS-6;复水后, ZSS结缕草及其新品系的颜色都有所恢复, 整体呈现ZSS-5≈ZSS-6>ZSS, 且新品系ZSS-5的叶片颜色一直处于深绿的状态.ZSS结缕草及其新品系的盖度随着胁迫的加剧而逐渐降低, 在胁迫第15天时, 盖度呈现ZSS-5>ZSS>ZSS-6;复水后, ZSS结缕草及其新品系的盖度都有所增加, 且新品系ZSS-5的盖度一直维持在100%.
|
|
表 3 干旱胁迫下ZSS-SJZ结缕草及新品系颜色得分和盖度的变化1) Table 3 Changes of color score and coverage of Zoysia japonica 'ZSS' and its new strains under drought stress |
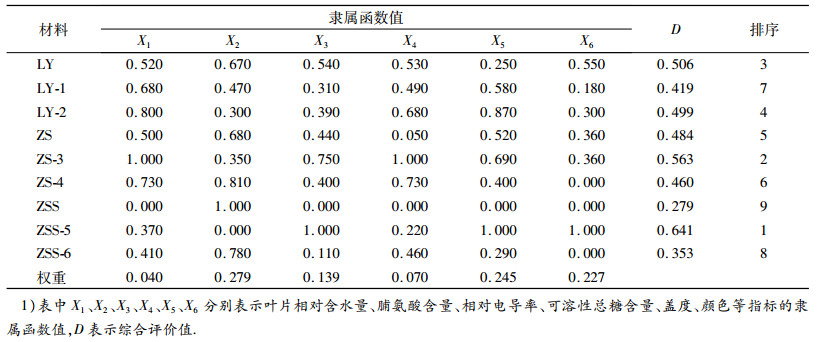
采用标准差系数赋予权重法, 选用叶片相对含水量、脯氨酸含量、相对电导率、可溶性总糖含量、盖度和颜色共6项指标, 对3种结缕草及其6种新品系材料的抗旱性进行了综合评价.综合评价值(D)为各种质材料在干旱胁迫下的抗旱性综合评价值, D值越大, 表明抗旱性越强.由表 4可以看出, 3种结缕草及其新品系抗旱性从强到弱依次为ZSS-5>ZS-3>LY>LY-2>ZS>ZS-4>LY-1>ZSS-6>ZSS.
|
|
表 4 干旱胁迫条件下结缕草隶属函数值1) Table 4 Subordinate function values of zoysiagrasses under drought stress |
RWC是植物抗旱分析的重要生理指标之一, 可反映植物水分亏缺程度, RWC的大小可部分反映植物的抗旱能力.在水分胁迫下, RWC减少得越多, 则草坪草所受的干旱伤害越大, 该草种的抗旱能力就越弱[15].本研究结果表明, 随着干旱胁迫程度的增加, 9种材料叶片相对含水量均有所下降.在干旱胁迫的第15天, 新品系ZSS-5和ZS-3的REC下降幅度小于其亲本, 表明在相同干旱胁迫下, 新品系植物对水分的利用效率更高, 叶片的保水能力更好, 说明其有更好的抗旱性.试验测定结果显示, LY-2和ZS-4的RWC较高, 表明其叶片保水能力相对较强, 但该结果与综合评价结果并不是完全一致, 说明用单一指标评价抗旱性可能导致片面性.
细胞原生质膜对干旱胁迫非常敏感, 干旱胁迫下植物细胞由于脱水伤害导致质膜相对透性增大, 从而使细胞内的电解质外渗, 导致电导率增大[16], 因此REC的高低可直接用来衡量草坪草的抗旱能力.在相同干旱胁迫下, 质膜透性增大的速度越大, 说明植物的抗旱性越弱[17].本研究结果表明, 在干旱胁迫的第15天, ZS-3和ZSS-5的REC显著低于各自亲本对照, 表明二者具有较强的抗旱性.
植物在严重干旱状态下, Pro和TSC作为渗透调节物质在植物体内的合成会增加[18-19].周兴元等[20]研究指出可将脯氨酸含量作为评价植物抗旱性的指标之一.余光辉[21]在假俭草Eremochloa ophiuriodes (Munro) Hack上的研究结果验证了这一结论.抗旱性弱的草种游离脯氨酸积累更为敏感, 积累的量比抗旱性强的草种多[22].本研究结果表明, 在整个干旱时期, 所有材料的Pro和TSC含量整体呈增大的趋势, 其中ZS-3和ZSS-5的Pro和TSC含量均显著低于各自亲本对照, 体内积累的含量也较低, 说明其具有较强的抗旱性.Cai等[23]认为, 抗旱性强的品种脯氨酸累积增多.上述2种截然相反的观点可能的原因是, 在干旱胁迫条件下不同植物对Pro和TSC敏感性不同, 因而存在调节程度的差别.
干旱胁迫下, 植物叶片的适应性变化将有利于水分的保持和水分利用效率的提高[24].通常采用草坪的综合评分来衡量茎叶的形态变化, 可以较为直观地说明草种的抗旱性[25].本研究结果表明, 在干旱胁迫的第9天, 土壤含水量(w)平均为12.74%, 所有的结缕草材料均能正常生长, 叶片色泽正常, 盖度保持100%;但在干旱胁迫的第15天时, 土壤含水量平均为2.43%, 大部分材料地上部生长减缓, 叶片枯黄、卷曲或对折, 盖度减小, 这与梁国玲等[3]的研究结果相似.干旱胁迫的第15天, LY-2、ZS-3、ZSS-5的盖度高于其亲本, 萎蔫率低于亲本, 并且复水之后恢复效果也较好, 表明这些新品系对干旱胁迫的适应能力较强.
植物的抗旱性由多基因控制, 是由多种因素相互作用而构成的一个较为复杂的综合性状.其中每一个因素与抗旱性本质之间存在着一定的联系或相关[26].单一的抗旱指标不能对植物的抗旱性做出评价, 需从整体上将生理指标和形态指标综合起来进行评价才能得出结论[27].本研究选用3种结缕草和经过60Co-γ辐射诱变之后筛选的6种新品系为材料, 通过对叶片的相对含水量、相对电导率、脯氨酸含量、可溶性总糖含量、盖度和颜色等指标的测定, 利用隶属函数加权法对9种材料的抗旱性进行了综合评价, 得出抗旱性从强到弱依次为ZSS-5>ZS-3>LY>LY-2>ZS>ZS-4>LY-1>ZSS-6>ZSS.说明结缕草可通过辐射育种选育出抗旱品种, 这为草坪草抗旱育种奠定基础.
| [1] |
WANG W X, VINOCUR B, ALTMAN A. Plant responses to drought, salinity and extreme temperatures:Towards genetic engineering for stress tolerance[J]. Planta, 2003, 218(1): 1-14. DOI:10.1007/s00425-003-1105-5 (  0) 0) |
| [2] |
徐炳成, 山仑. 草坪草对干旱胁迫的反应及适应性研究进展[J]. 中国草地, 2001, 23(2): 55-61. ( 0) |
| [3] |
梁国玲, 周青平, 颜红波, 等. 羊茅属4种牧草苗期抗旱性鉴定[J]. 草地学报, 2009, 17(2): 206-212. ( 0) |
| [4] |
WHITE R H, ENGELKE M C, MORTON S T, et al. Irrigation water requirement of zoysiagrass[J]. Int Turfgrass Soc Res J, 1993, 7: 587-593. ( 0) |
| [5] |
胡化广, 刘建秀, 宣继萍, 等. 结缕草属植物的抗旱性初步评价[J]. 草业学报, 2007, 16(1): 47-51. DOI:10.3321/j.issn:1004-5759.2007.01.008 ( 0) |
| [6] |
HUANG B, DUNCAN R R, CARROW R N. Drought-resistance mechanisms of seven warm-season turfgrasses under surface soil drying:Ⅱ:Root aspects[J]. Crop Sci, 1997, 37(6): 1863-1869. DOI:10.2135/cropsci1997.0011183X003700060033x ( 0) |
| [7] |
吕静, 刘卫东, 王丽, 等. 4种暖季型草坪草的抗旱性分析[J]. 中南林业科技大学学报, 2010, 30(3): 100-104. DOI:10.3969/j.issn.1673-923X.2010.03.017 ( 0) |
| [8] |
宣继萍. 结缕草属(Zoysia Willd. )植物种质资源多样性研究[D]. 南京: 南京农业大学, 2008.
( 0) |
| [9] |
JOONKI K, INJA S, SEOYOUNG S, et al. Production of unbolting lines through gamma-ray irradiation mutagenesis in genetically modified herbicide-tolerant Zoysia japonica[J]. Breeding Sci, 2009, 59(1): 103-105. DOI:10.1270/jsbbs.59.103 ( 0) |
| [10] |
余晓华, 张巨明, 王明祖, 等. 四种结缕草属草坪草对土壤干旱胁迫的响应及抗旱性研究[J]. 北方园艺, 2008(5): 121-124. ( 0) |
| [11] |
谢新春, 解彦峰, 张巨明, 等. 干旱胁迫下3种结缕草的抗旱性分析[J]. 广东农业科学, 2014, 41(17): 38-41. DOI:10.3969/j.issn.1004-874X.2014.17.010 ( 0) |
| [12] |
高俊风. 植物生理学实验指导[M]. 北京: 高等教育出版社, 2006, 15.
( 0) |
| [13] |
王学奎. 植物生理生化实验原理和技术[M]. 北京: 高等教育出版社, 2010, 174-176.
( 0) |
| [14] |
张荟荟, 甄世财, 张一弓, 等. 12份苜蓿种质材料苗期抗旱性综合评价[J]. 草业科学, 2014, 31(4): 737-742. ( 0) |
| [15] |
胡化广, 张振铭, 郭敏. 四种草坪草对干旱胁迫的反应及抗旱性评价[J]. 节水灌溉, 2011(11): 37-40. ( 0) |
| [16] |
BLUM A, EBERCON A. Cell membrane stability as measure of drought and heat tolerance in wheat[J]. Crop Sci, 1980, 21(1): 43-47. ( 0) |
| [17] |
LARKINDALE J, HUANG B. Changes of lipid composition and saturation level in leaves and roots for heatstressed and heat-acclimated creeping bentgrass[J]. Environ Exp Bot, 2004, 51(1): 57-67. DOI:10.1016/S0098-8472(03)00060-1 ( 0) |
| [18] |
刘刊, 耿士均, 商海燕, 等. 草坪草抗性研究进展[J]. 草业科学, 2012, 29(7): 7. ( 0) |
| [19] |
马芳蕾. 芒属五种观赏草的抗旱性研究[D]. 福州: 福建农林大学, 2014.
( 0) |
| [20] |
周兴元, 曹福亮, 刘国华. 两种暖季型草坪禾草对土壤持续干旱胁迫的生理反应[J]. 草业学报, 2004, 13(1): 84-88. DOI:10.3321/j.issn:1004-5759.2004.01.015 ( 0) |
| [21] |
余光辉. 水分胁迫下假俭草脯氨酸累积的ABA、Ca2+调节[D]. 广州: 华南师范大学, 2003.
( 0) |
| [22] |
田福平, 路远, 张小甫, 等. 苜蓿新品种(系)的抗旱性综合评价[J]. 江苏农业科学, 2014, 42(1): 160-163. DOI:10.3969/j.issn.1002-1302.2014.01.059 ( 0) |
| [23] |
CAI H, WU F, YANG W. Effect of simulated drought stress on antioxidant enzyme system of four native species in the ecotone between the Minjiang dry valley and mountain forest[J]. Chin J Appl Environ Biol, 2010, 16(4): 478-482. ( 0) |
| [24] |
刘颖, 张巨明. 暖季型草坪草对干旱胁迫的反应[J]. 草业科学, 2013, 30(11): 1732-1738. ( 0) |
| [25] |
JIANG Y, HUANG B. Drought and heat stress injury to two cool-season turfgrasses in relation to antioxidant metabolism and lipid peroxidation[J]. Crop Sci, 2001, 41(2): 436-442. DOI:10.2135/cropsci2001.412436x ( 0) |
| [26] |
降云峰, 赵晋锋, 马宏斌, 等. 作物干旱研究进展[J]. 中国农学通报, 2013, 29(3): 1-10. ( 0) |
| [27] |
王志泰, 马瑞, 马彦军, 等. 利用隶属函数法分析胡枝子抗旱性[J]. 干旱区资源与环境, 2013, 27(9): 121-123. ( 0) |