 2015, Vol. 36
2015, Vol. 36



2. 中国科学院 华南植物园,广东 广州 510650
2. South China Botanical Garden, Chinese Academy of Sciences, Guangzhou 510650, China
桉树Eucalyptus spp.是桃金娘科Myrtaceae桉属的总称.因其耐干旱瘠薄、适应性广、速生高产等特点,被联合国粮农组织推荐为三大速生造林树种之一[1],同时也是理想的生物原料[2].桉树人工林的生态退化,破坏了区域水分平衡[3-4],需大量施肥来维持桉树人工林高产[5].但于福科等[6]认为桉树人工林生产力下降实际上是林地土壤质量退化和生物多样性减弱的后续效应与必然结果,桉树耗水对于生态环境的影响仍存在争议.树木蒸腾耗水的测量方法经过了3个阶段[7].早期主要是快速称质量法.迅速发展阶段有蒸渗仪法、光合系统测定仪法等[8-9].到了20世纪90年代,热扩散法、放射性同位素以及能量平衡等测定方法出现[10],其中快速称质量法仍为目前最常用的方法[11].巨桉E. grandis和尾巨桉E. urophylla × E. grandis由于适应性强、速生、丰产、优质等特点而被广泛种植利用.胡红玲等[12]认为巨桉具有较高的耗水能力[13].尾巨桉的日平均蒸腾速率变化规律近单峰曲线[14].尾巨桉和巨桉作为目前华南地区栽种的主要桉树人工林树种,二者耗水间的差异存在较大争议,类似研究也相对较少.
对植物耗水特性进行深入研究,估测不同植物单位面积的蒸腾量,准确地为苗木进行浇水灌溉,对解决水资源利用与节水的矛盾具有重要意义,从而也为苗木进行高效定向培育提供了理论依据和实践指导.其中,蒸腾耗水速率和水分利用效率是树木重要的抗旱指标[15]和干旱适应性指标[16].本文通过对2种尾巨桉无性系(DH33-27和DH32-29)和巨桉无性系(H1)的耗水特性研究,评价其在不同土壤水分条件下的水分消耗特征和耗水能力差异,为正确评价这3个速生树种水分消耗和利用特点提供理论支持,并为科学审视桉树人工林水资源消费问题提供参考.
1 材料与方法 1.1 试验材料试验材料为来自南方国家级林木种苗示范基地的生长优良的2种尾巨桉无性系(DH33-27和DH32-29)和巨桉无性系(H1). 3种苗木均为1年生组培苗. 2012年12月中旬将所有试验材料上盆,每盆1株.花盆规格为200 mm × 150 mm,基质土壤为泥炭土和黄土的混合土,其中泥炭土由市场购入,黄土取自华南农业大学树木园,其混合比例为m(黄土): m(泥炭土)= 7: 3,每盆土的质量为(4.0 ± 0.2)kg.定期浇水、除草、防病虫害,以保证苗木正常生长. 2013年6月份试验前苗木生长情况如表 1所示.
|
|
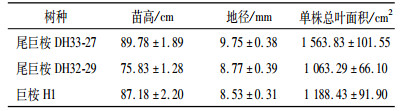
表 1 3种苗木试验前基本情况 Table 1 Growth status for three seedlings |
采用盆栽苗木称质量法对苗木的耗水特性进行测定. 2013年6月25日将试验苗木浇透水,每种选择6株生长正常、长势(地径和苗高)接近的苗木设为重复,将苗盆用塑料袋完全套住(从苗木根茎处覆盖整个表面,并密封花盆底部),防止土壤水分的蒸发.每次称量在08:00、10:00、12:00、14:00、16:00、18:00、20:00和翌日08:00分别用SP-30电子天平(美国)称盆质量1次,同时,用温湿度计(美德时TH603A)记录每次称盆时环境空气的温度和湿度.叶面积计算与邱权等[17]一致,根据公式(分级叶片数×叶面积再相加之总和)得到单株叶面积.根据盆质量称量结果分别计算出白天、夜晚以及白天每个时间段(2 h)的耗水量,并用公式计算出对应的耗水速率:耗水速率=每个时间段的耗水量/(单株叶面积×时间).
1.2.2 土壤水分条件设置由于不同的水分胁迫程度还没有统一的划分标准,因此一般用土壤含水量来确定植株的干旱程度[18].本试验中,利用FOM /mts便携式土壤湿度计对试验期土壤体积含水量进行测定.每树种按照测出的体积含水量分为4个等级:封盆后0 ~ 3 d视为处于正常水分条件时期,作为对照(CK),体积含水量≥18%;封盆后4 ~ 6 d视为轻度干旱(Light dry,LD)时期,体积含水量为13% ~ 18%;封盆后7 ~ 15 d视为中度干旱(Medium dry,MD)时期,体积含水量为9% ~ 13%;封盆后16 ~ 23 d视为严重干旱(Severe dry,SD)时期,体积含水量低于9%.
1.2.3 叶片水分利用效率测定2013年6月,用Li-6400便携式光合作用分析系统(美国),选择3个晴天,于09:30-11:00测定苗木叶片净光合速率(Pn)和蒸腾速率(Tr),每种苗木选择3株,每株测定3片,其中,光照度设置为1 000 μmol·m-2·s-1,温度35 ℃.水分利用效率(WUE)= Pn / Tr[19].
1.2.4 数据处理采用Excel 2007进行作图,SPSS对试验数据进行方差分析、多重比较(Duncan′s法)和相关性分析.
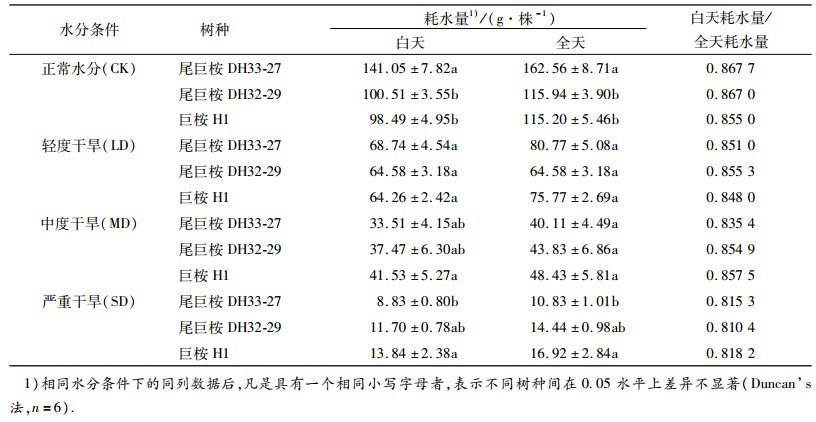
2 结果与分析 2.1 单株苗木耗水量的比较从表 2可以看出,尾巨桉DH33-27在CK时期白天耗水量最大[(141.05 ± 7.82)g·株-1],显著高于其他2种树种.在LD和MD时期,下降幅度较大,在SD时期白天耗水量最低,仅(8.83 ± 1.01)g·株-1,显著低于巨桉H1(13.84 ± 2.38)g·株-1(P<0.05),但尾巨桉DH32-29与二者差异不显著.在处理过程中,尾巨桉DH32-29的苗木耗水量仅在CK时期与尾巨桉DH33-27出现了显著差异(P<0.05).尾巨桉DH32-29各时期耗水量介于尾巨桉DH33-27与巨桉H1之间.巨桉H1在4个时期耗水量变化最小,SD时期的全天耗水量占CK时期的14.69%,而尾巨桉DH33-27则为6.66%. 3种树种的白天耗水量均占全天的80%以上,但随着干旱程度的不断加深,比值有所下降,从CK时期的85.50% ~ 86.77%下降到SD时期的81.04% ~ 81.82%.
|
|
表 2 不同水分条件下苗木平均耗水量 Table 2 The average water consumption of seedlings under different soil moisture contents |
由图 1可见,每种树种白天最大耗水速率出现在10:00-14:00,同时耗水速率呈“单峰”曲线变化.在正常土壤水分条件下,尾巨桉DH33-27和DH32- 29白天各时段耗水速率均大于巨桉H1,且巨桉H1白天最大耗水速率(94.80 ± 4.54)g·h-1 ·m-2出现在10:00-12:00时段. 3种树种在正常水分条件下最大耗水速率与白天平均耗水速率的比值分别为1.31(尾巨桉DH33-27)、1.39(尾巨桉DH32-29)和1.43(巨桉H1). LD时期,3种树种最大耗水速率均出现于10:00-12:00,白天各时段耗水速率曲线相似.在MD时期,尾巨桉DH33-27和DH32-29白天平均耗水速率剧烈下降,明显低于巨桉H1(44.50 ± 2.09)g·h-1·m-2.随着水分胁迫日益严重至苗木死亡前期,每种树种的白天各时段耗水速率曲线慢慢趋向于平缓且规律不明显,在SD时期,尾巨桉DH33-27、尾巨桉DH32-29和巨桉H1日平均耗水量相对于CK时期分别减少了92.34%、87.83%和82.71%.由图 2可得,随着干旱胁迫程度加深,苗木白天平均耗水速率均明显下降,且尾巨桉DH33-27和巨桉H1呈现显著性差异.

|
图 1 不同土壤水分条件下苗木白天各时段平均耗水速率 Figure 1 The average water consumption rate of seedlings in each period in the daytime under different soil moisture contents |

|
图 2 不同土壤水分条件下苗木白天平均耗水速率 Figure 2 The average water consumption rate of seedlings during daytime under different soil moisture contents 相同水分条件下的柱子上方,凡是具有一个相同小写字母者,表示不同树种间在0.05水平上差异不显著(Duncan’s法,n = 6). |
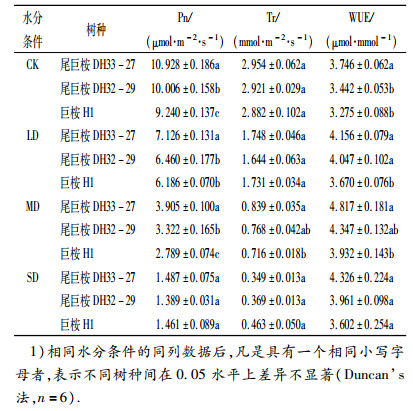
通过对3种树种的Pn、Tr和WUE进行相互比较进一步分析桉树苗木的水分利用效率.由表 3可以看出,在正常水分条件下,3种树种的Pn存在显著性差异(P<0.05),Tr不存在显著性差异,而尾巨桉DH33- 27的WUE与其余2种树种存在显著性差异.随着干旱胁迫程度的加重,在LD和MD时期,Pn和Tr明显下降,WUE上升.在SD时期,尾巨桉DH33-27、DH32- 29和巨桉H1的Pn与Tr持续下降,而WUE在MD时期出现最高值后有所回落,在SD时期为3.602 ~ 4.326 μmol·mmol-1.水分利用效率由大到小依次为:尾巨桉DH33-27>尾巨桉DH32-29>巨桉H1.
|
|
表 3 3种苗木水分利用效率的比较1) Table 3 A comparison of water use efficiency for three kinds of seedlings |
苗木耗水量是反映树种耗水潜力最直接的指标[15].在7月份高温低湿的环境中,从树种全天耗水量的变化可以初步判断树种的节水性能.但是,要全面了解不同树种间耗水能力的差异,仅凭耗水量不够,还应结合单位时间树种在单位叶面积的耗水量,即耗水速率来评价才更为准确.先前研究证实巨桉H1为高耗水量树种[13],而本研究中,在水分充足的条件下,巨桉H1总耗水量(115.20 ± 5.46)g·株-1与尾巨桉DH32-29(115.94 ± 3.90)g·株-1十分接近,并且远小于尾巨桉DH33-27(162.56 ± 8.71)g·株-1,到了SD时期其总耗水量(16.92 ± 2.84)g·株-1则远高于二者,且存在显著性差异.说明巨桉H1较2种尾巨桉具有更差的节水能力,表现为在受到干旱胁迫时仍然保持较高的耗水量.这与胡红玲等[12]认为的巨桉不仅具有较高的耗水量,且在一定的干旱程度内,也更不易受到干旱影响而降低耗水量的观点一致.而尾巨桉在干旱后期耗水剧烈下降是否与其叶片大量掉落造成总叶面积减少有关,还需更多试验数据加以证明.当然,树木蒸腾作用是由气孔导度和边界层导度共同控制的[20-22],当干旱胁迫加重,水分便主要通过皮孔蒸腾或角质层蒸腾来减少[23].本研究中,随着水分胁迫的加剧,苗木保卫细胞缺水,控制气孔内外渗透压,导致气孔阻力剧增,也是蒸腾耗水减少的重要原因.这是否还与叶脉致密度、叶片上蜡面沉积面积相关还需更深入研究.
在众多关于苗木蒸腾耗水的研究中,学者们均得出相似的结论,即正常水分条件下最大耗水速率与白天平均耗水速率的比值为1.6[24-26],在本研究中,该比值为1.31 ~ 1.43,与邱权等[17]对巨桉的研究结果一致.最大耗水速率与白天平均耗水速率的比值可推测苗木对气候的敏感程度,比值越高可能说明叶片敏感度越高,受到环境因素影响越迅速,却似乎不能证明树种的节水能力强弱,但这一稳定的比值对瞬时蒸腾速率推算耗水量有重要的意义.
3.2 桉树间水分利用效率比较植物耗水量与叶面积和叶片蒸腾强度有关[27].在上述分析中发现尾巨桉DH32-29耗水量接近于巨桉H1,与CK时期二者WUE不存在显著性差异的结论一致.本研究数据表明:与其余2种桉树相比,巨桉H1在干旱胁迫过程中一直保持较低的水分利用效率,同时,光合速率和蒸腾速率也较尾巨桉DH33- 27低.在CK、LD和MD时期,3种树种的WUE逐步提高,在SD时期有所下降,这是由于树种受到干旱胁迫后,植物为了维持生命活动,吸收的水分更多地被用于光合作用而不是通过蒸腾散失,水分利用效率短暂提高正是苗木适应干旱的表现.死亡前期3种树种水分利用效率并无显著性差异,说明植物各机能此时已降到了最低,并且供试树种均来自桉属,推测其具有相似的干旱适应能力.综上所述,相比尾巨桉DH33-29和巨桉H1,尾巨桉DH33-27是高光合、高水分利用效率的品系,不仅适宜在水分条件好的地区大面积推广,也适宜在水分严重亏缺的地区种植.树木生长要遵循“适地适树”的原则,不仅要考虑当地的气候状况,也应从水分条件进行思考.此次试验为盆栽试验,能否在人工林中也表现出同样特性,不仅需要考虑不同苗木耗水特性和水分利用效率,也应综合考虑各方面因素比如当地水资源、气候变化、经纬度等.
3.3 与其他速生树种耗水特性的对比植物的抗旱能力是一个复杂的特征,以楸树为例[28],影响最大的4个指标为:叶片厚度、栅栏组织厚度、气孔密度以及叶片游离脯氨酸变化量.耗水量虽然能直观反映不同树种的节水能力,但由于受到叶面积影响较大,所以一般都以耗水速率和水分利用效率为主要参考指标.耗水速率是植物固有的生理特性,稳定性好,能反映植物调节自身水分耗损能力和在不同环境中的实际耗水特征,因此常用来比较不同植物固有的耗水能力[29].为了更好地对尾叶桉、巨桉与其他速生树种的耗水性能进行比较,特将此3种树种与尾叶桉(广林- 9号)、楸树和毛白杨进行对比.其数据测定时间均为当年7月份,测定方法均为盆栽苗木称质量法,一年生苗木,因此具有较大的可比性.
由何茜[25]试验得知:毛白杨Populus tomentosa的平均白天耗水量为每株177.30 g·d-1,平均耗水速率为90.00 g·m-2·h-1.尾巨桉(广林-9号)的平均白天耗水量为每株162.07 g·d-1,平均耗水速率为48.23 g·m-2·h-1;竹柳(Bamboo-willow)平均白天耗水量为每株88.78 g· d-1,平均耗水速率为59.19 g·m-2·h-1 [17].正常水分条件下,这6种树种耗水能力差异较大,同样是尾巨桉的3个不同品种也存在较大差异.这6种树种耗水速率大小排名为毛白杨>尾巨桉DH32-29>尾巨桉DH33-27>巨桉H1>竹柳>尾巨桉(广林- 9号).因此,从单株苗木耗水角度来讲,认为桉树高耗水缺乏科学性,与华雷等[30]试验结果相同.比较不同树种在充足的水分条件下耗水能力,不仅应比较不同树种间的耗水量,也应考虑其耗水速率.综上所述,为了充分利用稀缺水资源,为苗木生长进行因地制宜的充足灌溉,研究苗木耗水特性十分必要.本研究证明,尾巨桉、巨桉都是节水性能较好的树种,在对速生树种进行人工造林时可重点考虑.
| [1] |
杨民胜, 吴志华, 陈少雄, 等. 桉树的生态效益及其生态林经营[J]. 桉树科技, 2006, 23(1): 32-39. DOI:10.3969/j.issn.1674-3172.2006.01.007 (  0) 0) |
| [2] |
BRITTO D S, PIROVANI C P, GONZALEZ E R, et al. Oxidative stress proteins as an indicator of a low quality of eucalyptus clones for the pulp and paper industry[J]. Genet Mol Res, 2012, 11(4): 3798-3813. DOI:10.4238/2012.August.17.13 ( 0) |
| [3] |
YU F, HUANG X, DUAN C, et al. Eucalyptus trees-Ageratina adenophora complex system: A new eco-environmental protection model[J]. Clean-Soil Air Water, 2014, 42(5): 682-689. DOI:10.1002/clen.v42.5 ( 0) |
| [4] |
ESHETU Y, OLAVI L. Indigenous woody species diversity in Eucalyptus globulus Labill. ssp. globulus plantations in the Ethiopian highlands[J]. Biodivers Conserv, 2003, 12(3): 567-582. DOI:10.1023/A:1022483700992 ( 0) |
| [5] |
陈少雄. 桉树人工林土壤养分现状与施肥研究[J]. 桉树科技, 2009, 26(1): 52-63. DOI:10.3969/j.issn.1674-3172.2009.01.011 ( 0) |
| [6] |
于福科, 黄新会, 王克勤, 等. 桉树人工林生态退化与恢复研究进展[J]. 中国生态农业学报, 2009, 17(2): 393-398. ( 0) |
| [7] |
孙鹏森, 马履一. 水源保护树种耗水特性研究与应用[M]. 北京: 中国环境科学出版社, 2002.
( 0) |
| [8] |
苏建平, 康博文. 我国树木蒸腾耗水研究进展[J]. 水土保持研究, 2004, 11(2): 177-179. DOI:10.3969/j.issn.1005-3409.2004.02.057 ( 0) |
| [9] |
郭孟霞, 毕华兴, 刘鑫, 等. 树木蒸腾耗水研究进展[J]. 中国水土保持科学, 2006, 4(4): 114-120. DOI:10.3969/j.issn.1672-3007.2006.04.022 ( 0) |
| [10] |
王颖, 余瑞卿, 李湛东, 等. 城市片林中常见树种的蒸腾耗水特性研究综述[J]. 内蒙古农业大学学报:自然科学版, 2005, 26(3): 115-119. ( 0) |
| [11] |
茹桃勤, 李吉跃, 孔令省, 等. 刺槐耗水研究进展[J]. 水土保持研究, 2005, 12(2): 135-140. DOI:10.3969/j.issn.1005-3409.2005.02.042 ( 0) |
| [12] |
胡红玲, 张健, 胡庭兴, 等. 不同土壤水分条件下三个速生用材树种幼树的耗水特性比较[J]. 自然资源学报, 2012, 27(12): 2071-2080. DOI:10.11849/zrzyxb.2012.12.008 ( 0) |
| [13] |
胡红玲. 巨桉耗水特性及其对干旱胁迫的响应[D]. 雅安: 四川农业大学, 2012.
( 0) |
| [14] |
卢春英. 闽西尾巨桉造林及其生长效果分析[J]. 福建林业科技, 2005, 32(2): 21-26. DOI:10.3969/j.issn.1002-7351.2005.02.006 ( 0) |
| [15] |
罗青红, 李志军. 树木水分生理生态特性及抗旱性研究进展[J]. 塔里木大学学报, 2005, 17(2): 29-33. DOI:10.3969/j.issn.1009-0568.2005.02.011 ( 0) |
| [16] |
AZAM G, MURRAY R S, GRANT C D, et al. Tolerance of young seedlings of Different tree species and a cereal to poor soil aeration[J]. Soil Res, 2014, 52(8): 751-759. DOI:10.1071/SR13219 ( 0) |
| [17] |
邱权, 潘昕, 李吉跃, 等. 速生树种尾巨桉和竹柳幼苗耗水特性和水分利用效率[J]. 生态学报, 2014, 34(6): 1401-1410. ( 0) |
| [18] |
张雪海, 李吉跃, 刘娟娟, 等. CO2倍增与干旱胁迫交互作用对幼树蒸腾耗水日变化的影响[J]. 广东林业科技, 2009, 25(2): 1-9. DOI:10.3969/j.issn.1006-4427.2009.02.001 ( 0) |
| [19] |
PENUELAS J, FILELLA I, Llusià J, et al. Comparative field study of spring and summer leaf gas exchange and photobiology of the mediterranean trees Quercus ilex and Phillyrea latifolia[J]. J Exp Bot, 1998, 49(319): 229-238. ( 0) |
| [20] |
SALIENDRA N Z, SPETTY J S, COMSTOCK J P. Influence of leaf water status on stomatal responses to humidity, hydraulic conductance and soil drought in Betula occideratalis[J]. Planta, 1995, 196(2): 357-366. ( 0) |
| [21] |
HIROMI T, NINOMIYA I, KOIKE T, et al. Regulation of transpiration by patchy stomatal opening in canopy tree species of Dipterocarpaceae in tropical rain forest, Sarawak, Malaysia[J]. Jpn J Ecol, 1999, 49(2): 83-90. ( 0) |
| [22] |
奚如春, 马履一, 王瑞辉, 等. 林木耗水调控机理研究进展[J]. 生态学杂志, 2006, 25(6): 692-697. DOI:10.3321/j.issn:1000-4890.2006.06.020 ( 0) |
| [23] |
李吉跃, 朱妍. 干旱胁迫对北京城市绿化树种耗水特性的影响[J]. 北京林业大学学报, 2006, 28(S1): 32-37. ( 0) |
| [24] |
朱妍, 李吉跃, 史剑波. 北京六个绿化树种盆栽蒸腾耗水量的比较研究[J]. 北京林业大学学报, 2006, 28(1): 65-70. DOI:10.3321/j.issn:1000-1522.2006.01.012 ( 0) |
| [25] |
何茜. 毛白杨抗旱节水优良无性系评价与筛选[D]. 北京: 北京林业大学, 2008. http://cdmd.cnki.com.cn/Article/CDMD-10022-2008085056.htm
( 0) |
| [26] |
陈博. 珍贵树种楸树优良无性系评价与筛选[D]. 广州: 华南农业大学, 2012.
( 0) |
| [27] |
马履一, 王瑞辉, 徐军亮, 等. 北京市主要园林绿化植物耗水性及节水灌溉制度研究[M]. 北京: 中国林业出版社, 2009, 5.
( 0) |
| [28] |
董蕾, 陈博, 李吉跃, 等. 5个楸树无性系抗旱性的综合评价[J]. 中南林业科技大学学报, 2014, 34(4): 30-35. DOI:10.3969/j.issn.1673-923X.2014.04.007 ( 0) |
| [29] |
邱权, 何茜, 李吉跃, 等. 干旱胁迫对华南地区三种苗木蒸腾耗水的影响[J]. 广东林业科技, 2012, 28(1): 19-24. DOI:10.3969/j.issn.1006-4427.2012.01.004 ( 0) |
| [30] |
华雷, 何茜, 李吉跃, 等. 桉树无性系和华南乡土树种秋枫苗木耗水特性的比较[J]. 应用生态学报, 2014, 25(6): 1639-1644. ( 0) |