 2016, Vol. 37
2016, Vol. 37



2. 华南农业大学 林学与风景园林学院/亚热带农业生物资源保护与利用国家重点实验室/广东省森林植物种质创新与利用重点实验室,广东 广州 510642;
3. 广东省九连山林场,广东 河源 517100
2. College of Forestry and Landscape Architecture, South China Agricultural University/ State Key Laboratory for Conservation and Utilization of Subtropical Agro-bioresources/Guangdong Key Laboratory for Innovative Development and Utilization of Forest Plant Germplasm, Guangzhou 510642, China;
3. Jiulianshan Forest Farm of Guangdong, Heyuan 517100, China
黄梁木Neolamarckia cadamba,又称团花树,为我国南方乡土阔叶树种,由于生长迅速,树干通直,因而被誉为“奇迹树”,早在20世纪70年代就受到国内外普遍关注。其材性与杉木相当,速生性与桉树、杨树相近,是较好的用材和纸浆原料树种;树皮含有丰富的生物碱类物质,其中3α-二氢卡丹宾和3β-二氢卡丹宾为治疗高血压药物“钩藤总碱”的有效成分,具有强而持久的降压作用,其效价已经接近利血平[1],这2个生物碱类物质在钩藤Uncaria rhynchophylla中仅为微量成分存在,而在黄梁木树皮中作为仅次于卡丹宾含量的主要成分存在,是较好的药源树种。此外,黄梁木也是优良的园林绿化、蜜源和饲料树种[2]。黄梁木用途广,潜在经济价值大,有着广阔的开发前景。
因黄梁木有较好的速生、丰产特性,在云南、广西、广东和海南4省(区),人工种植面积已达270 hm2,造林农户和企业积极性较高,种植面积仍有增大的势头。国内外学者已对黄梁木的种子生理[3]和繁殖[4-7]、引种驯化[8-9]、病虫害[10-11]、药用和化学成分[12-14]、生物技术[7, 15-19]、材性[20]等进行了深入研究,从常规育种到分子育种及其基因功能研究等均取得较好的结果,但抗寒性相关的研究鲜见报道。本文以云南、广西和广东共7个1.5年生黄梁木种源的带芽枝条为材料,通过人工低温胁迫处理,采用隶属函数法和主成分分析法对不同种源抗寒能力进行综合评价,为黄梁木抗寒育种早期鉴定和引种栽培提供科学依据。
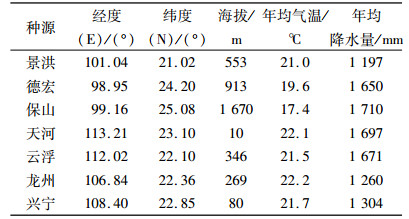
1 材料与方法 1.1 试验材料从1.5年生实生苗植株剪取长度约为20 cm的带芽枝条作为试验材料,景洪、德宏、保山为云南种源,天河、云浮为广东种源,龙州、兴宁为广西种源。各种源来源情况详见表 1。种子均采集于10年以上树龄大树;种子播种后,苗高约20 cm时移植到施有20 kg农家肥的种植穴内,按需浇水,1年后开始在根部追施复合肥。种植地位于广东省增城市华南农业大学科研基地,地理位置E 113°37′,N 23°14′,海拔20.3 m,平均气温21.8 ℃,年均降水量2 137 mm;土质为黏壤土,地势平坦。
|
|
表 1 黄梁木种源地理位置与气候因子 Table 1 Geographical position and climatic factors of Neolamarckia cadamba provenances |
据黄梁木适生地气象记录、实地调查受寒害情况和低温胁迫预试验,低温胁迫处理梯度设置为8、4、2、0、-2、-4、-6和-8 ℃。以不做低温胁迫处理的材料作为对照(CK)。
于2013年12月中下旬(气温12~15 ℃),随机剪取同一种源黄梁木植株中上部朝向和着生部位一致、长势旺、无病虫害的枝条,清洗表面脏物,吸干水分,每种源枝条剪取540根,分成6等份,作为每个生理指标测定和恢复生长材料,即每个处理30根,3次重复。其中,0 ℃以上低温不做预处理,直接置于人工气候箱中进行低温胁迫处理12 h;0 ℃及以下低温胁迫处理,先置于4 ℃下处理2 h,再将温度降至相应的低温处理12 h。在低温胁迫处理结束后,均在12 ℃中恢复12 h。试验处理中的升、降温速度均为2 ℃·h-1。
1.2.2 测定指标相对电导率(REC)、游离脯氨酸(用磺基水杨酸法)按王晶英[21]的方法测定,丙二醛(硫代巴比妥酸显色法)、可溶性糖含量(蒽酮比色法)、可溶性蛋白质含量(考马斯亮蓝G-250染色法)按李合生[22]的方法测定。
1.2.3 恢复生长试验将低温胁迫处理后的带芽枝条,于25 ℃、光照强度2 500~3 000 lx、光周期12 h的条件下水培,每2天更换1次清水(每周用含MS大量元素成分的营养液培养2 d),20 d后观察并记录枝芽萌发情况,统计存活率,以存活率高低作为衡量抗寒性强弱的指标[23-26]。以长出嫩叶或芽点无褐化作为存活标准。
1.3 数据处理与分析方法 1.3.1 半致死温度计算参照朱根海等[27]和盖钧镒[28]的方法,建立相对电导率拟合Logistic回归方程,计算半致死温度(LT50)。Logistic回归方程为Y=K/(1+ae-bt),其中,Y代表胁迫处理温度下测得的相对电导率,K代表最大电导率(最大值为100),a、b为方程参数,t代表低温胁迫处理温度。求该回归方程二阶、三阶导数,并令其为0,可得如下公式[29-31]:
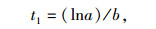
|
(1) |
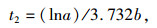
|
(2) |
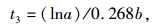
|
(3) |
其中, t1为回归曲线的拐点,即低温胁迫的半致死温度(LT50),t2为组织细胞膜受伤的起始温度,t3为组织细胞膜接近全透性时的温度。
按戴国俊等[32]、董江水[33]的方法,用SPSS 19.0软件估算方程参数,拟合方程,求出t1、t2和t3。
1.3.2 抗寒性综合指标计算用隶属函数法和主成分分析法进行综合评价[23, 25, 34-35]。
若主成分因子负荷量为正,采用升型分布函数,隶属函数公式为:
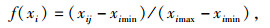
|
(4) |
若负荷量为负,则采用降型分布函数,隶属函数公式为:
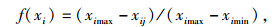
|
(5) |
其中,f(xi)为各指标抗寒隶属度值,xij为j种源第i项指标测定值,ximin、ximax分别为第i项指标的最小值、最大值。
据各指标隶属函数值与各指标权重,通过加乘法则对各指标进行合成,计算抗寒性综合指数。计算公式为:
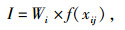
|
(6) |
式中,I为抗寒性综合指数,Wi为抗寒指标的权重,f(xij)为各指标的隶属函数值。最后通过I值大小对不同种源的抗寒能力强弱进行评价。
数据方差分析、多重比较、相关分析和聚类分析等均用SPSS 19.0软件完成。作图用Excel 2003软件完成。
2 结果与分析 2.1 不同种源枝芽在低温胁迫下相对电导率的变化及细胞膜受伤温度的确定从图 1可以看出,各种源在胁迫处理8~-8 ℃间,相对电导率的变化曲线基本呈“S”型,并且均在0~-4 ℃间变化较大。兴宁、云浮和天河种源在0~-4 ℃间的增加幅度略小,分别为28.16%、32.57%和34.06%,而保山和景宏种源的增加幅度较高,分别为46.43%和41.70%。在CK以及8~2 ℃低温间的变化曲线均近似于水平线,不同种源在相同低温处理下、同一种源在不同低温胁迫处理下的相对电导率变化较小;在胁迫温度小于-6 ℃后,各种源相对电导率变化减缓,并逐渐接近各自的最大值,表明细胞膜已接近完全受破坏。

|
图 1 黄梁木枝芽在不同低温胁迫处理下相对电导率的变化 Figure 1 The relative electrical conductivity changes in branch buds of Neolamarckia cadamba under low temperature stress |
相对电导率拟合的Logistic方程和细胞膜受伤温度见表 2。从表 2中可以看出,Logistic方程的决定系数R2为0.940~0.961,说明方程拟合度较好。不同种源的LT50为-0.74~-2.11 ℃,最高为德宏(-0.74 ℃),最低为云浮(-2.11 ℃)。云南3个种源的细胞膜受伤起始温度和死亡温度均为最高,分别为-0.20~-0.27和-2.78~-3.78 ℃;而广东、广西种源的细胞膜受伤起始温度和死亡温度明显低于云南种源:广东种源分别为-0.51~-0.57和-7.15~-7.89 ℃,广西种源分别为-0.49~-0.53和-6.83~-7.34 ℃。在所有种源中,云浮种源细胞膜受伤起始温度和死亡温度均最低,而德宏种源均最高。
|
|
表 2 7个不同种源拟合的Logistic方程和细胞膜受伤温度1) Table 2 The logistic equation and cell membrane injury temperature of seven provenances after treated with low temperature |
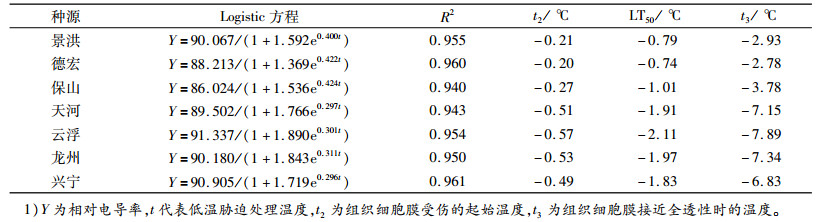
因此,从相对电导率拟合方程推导出的膜受伤温度看,抗寒性由弱到强的种源应依次为德宏、景洪、保山、兴宁、天河、龙州、云浮。
2.2 低温胁迫对不同种源恢复生长的影响不同低温胁迫处理下,黄梁木枝芽的存活率差异见表 3。当胁迫温度从0 ℃降至-2 ℃,7个种源存活率均下降明显:云南3个种源枝芽的存活率从几乎不受寒害迅速降低到4.0%左右(均值为4.15%),而广东和广西4个种源的枝芽存活率均从几乎不受寒害降低至30%~40%(均值分别为37.20%和36.38%)。在-2 ℃低温胁迫下,种源间存活率差异显著,景洪最低,为3.88%,云浮最高,为42.03%,广东和广西种源存活率均值分别是云南种源均值的8.96和8.76倍。当处理温度为-4 ℃时,云南3个种源几乎全部死亡,广东和广西4个种源仍有少量枝芽存活,云浮种源存活率最高,为8.17%;当处理温度为-6 ℃时,云南3个种源、龙州种源全部死亡,天河、兴宁和云浮种源均接近全部死亡,存活率分别为0.03%、0.02%和0.13%。这与用相对电导率拟合Logistic方程得出不同种源的半致死温度(LT50)在-0.74~-2.11 ℃基本一致。
|
|
表 3 不同低温处理后的枝芽存活率1) Table 3 Effects of low temperature treatments on branch buds survival rate |
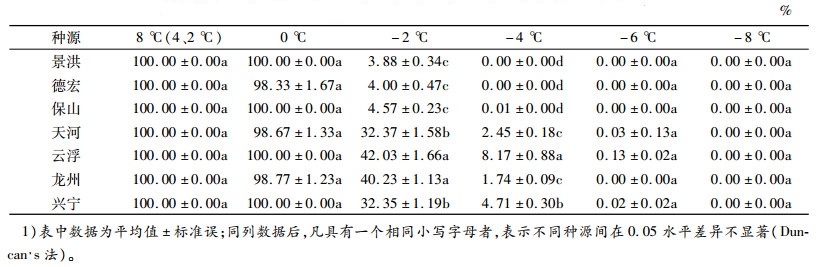
结果表明,各种源枝芽在低于0 ℃的胁迫温度下,存活率迅速下降,当胁迫温度降到-6 ℃时,几乎全部死亡;在不同胁迫温度下,云浮种源存活率均最高,表明云浮种源的抗寒能力最强。总体上看,广东种源的抗寒性较广西种源强,而云南种源最弱。
2.3 黄梁木抗寒性综合评价 2.3.1 抗寒综合评价中生理生化指标的选取植物遭受寒害后,评价植物受害程度或能否存活最直观的方法是观察枝芽恢复萌芽的能力[36]。从表 3可以看出,当胁迫温度低于-2 ℃后,枝芽的存活率极低,并且已有部分种源死亡。而经-2 ℃处理后,各种源枝芽的存活率均下降,并且种源间差异达显著水平,因此,可选取-2 ℃处理的枝芽存活率作为不同黄梁木种源抗寒性的衡量指标。在-2 ℃处理下的相对电导率、丙二醛含量、脯氨酸含量、可溶性糖含量、可溶性蛋白含量以及半致死温度见表 4。
|
|
表 4 7个种源在-2 ℃低温胁迫下的理化指标 Table 4 Physicochemical indicators of seven provenances under low temperature (-2 ℃) stress |
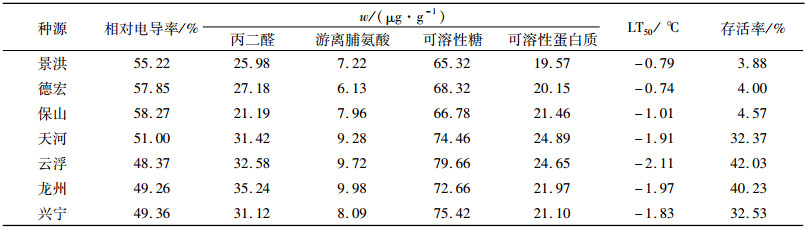
相关分析表明,枝芽存活率与丙二醛含量(0.897**)、游离脯氨酸含量(0.876**)、可溶性糖含量(0.912**)极显著正相关,与相对电导率(-0.964**)、半致死温度(-0.987**)极显著负相关,而与可溶性蛋白质(0.721)的相关性不显著。说明丙二醛含量、游离脯氨酸含量、可溶性糖含量、相对电导率、半致死温度对黄梁木抗寒性影响较大,而可溶性蛋白质对其抗寒性影响相对稍小。因此,选取与枝芽存活率极显著相关的丙二醛含量、游离脯氨酸含量、可溶性糖含量、相对电导率、半致死温度作为抗寒性综合评价的指标。
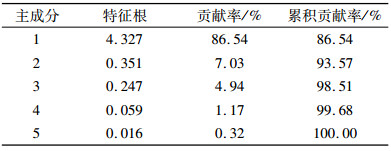
2.3.2 抗寒指标权重确定由于用单一生理和生化指标对抗寒性评价具有一定的片面性,而且不同生理生化指标之间的关系复杂,对抗寒性的重要性不同,因此,对选取的5个抗寒生理生化指标进行主成分分析。结果(表 5)表明,第1个主成分的特征根大于1,贡献率达86.54%,其他主成分的特征根均小于1,贡献率较小。故用第1个主成分就能够代表原5个生理生化指标变量的信息(即此主成分指标包含原5个抗寒指标86.54%的相关信息)。因此,以第1主成分分析各指标的负荷量,计算各指标对黄梁木抗寒性作用大小,确定权重Wi(表 6)。
|
|
表 5 抗寒性相关指标的特征根和贡献率 Table 5 The eigenvalue and contribution rate of cold resistance indicators |
|
|
表 6 抗寒性指标的负荷量和权重 Table 6 Capacities and weights of cold resistance indicators |
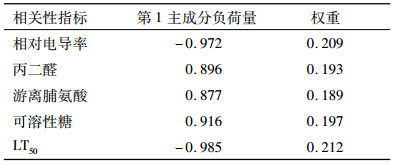
由表 6可见,对黄梁木抗寒性影响最大的指标是LT50,其次是相对电导率,然后依次为可溶性糖、丙二醛、游离脯氨酸。
2.3.3 抗寒指标的隶属度鉴于抗寒指标的单位、性质各不相同,但各指标的变化具有连续性,因此对这些指标进行标准数量化后,按主成分因子负荷量,采用隶属度函数法确定隶属函数的升降顺序[37]。丙二醛、游离脯氨酸、可溶性糖3个指标采用升型分布函数,相对电导率和LT50则采用降型分布函数。不同种源抗寒性指标隶属度值见表 7。
|
|
表 7 7个不同种源抗寒性指标的隶属度值 Table 7 Subordination values of cold resistance indicators of seven provenances |
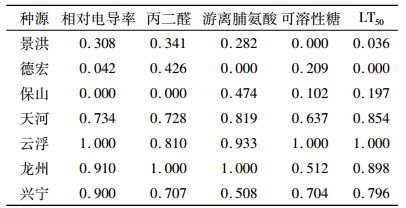
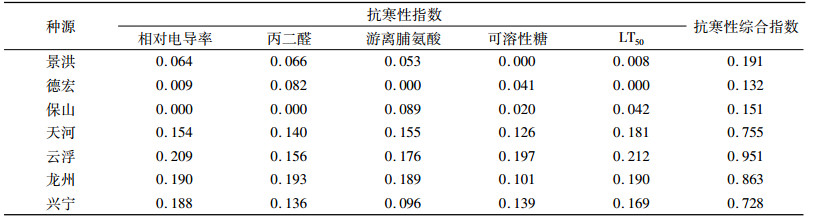
根据上述各指标的隶属度值和权重,通过式(6)计算抗寒性综合指数I(表 8),综合指数越大,表示抗寒性越强。由表 8可知,不同种源抗寒性从强到弱依次为云浮、龙州、天河、兴宁、景洪、保山、德宏。
|
|
表 8 7个不同种源抗寒性综合指数 Table 8 Integrated indices of cold resistance indicators of seven provenances |
不同种源黄梁木在低温胁迫处理后,相对电导率的变化趋势基本一致,均随着胁迫温度的降低呈“S”型曲线上升。相对电导率拟合的Logistic方程结果表明,试验种源的半致死温度在-0.74~-2.11 ℃间,云浮种源的半致死温度最低。种源间在-2 ℃低温处理下存活率差异显著,均在45%以下,表明大部分枝芽已受到严重的寒害,与2011年1月广东省新兴县的黄梁木人工林遭受-1.5 ℃低温造成枝芽和顶芽枯死的调查结果(另文发表)相近。
大量研究结果表明,植物抗寒生理变化过程是错综复杂的,孤立地用某一生理指标很难反映植物对寒冷的综合适应能力[38]。在小麦Triticum aestivum[39]、珍珠菜属Lysimachia植物[35]、葡萄Vitisvinifera[40-41]、蜡梅Chimonanthus praecox[42]、翅荚木Zeniainsignis[24]等的抗寒研究中已发现,利用单项抗寒生化或理化指标鉴定植物的抗寒性局限性很大。而在核桃Juglansregia[43]、扁桃Amygdaluscommunis[34]、早实品种的核桃[44]、平欧杂交榛Coryusheterophylla×Coryusavellan[23]等植物的抗寒性研究中,利用了多个抗寒生化或理化指标进行综合评价,获得较满意的效果。本文采用隶属函数法和主成分分析法,利用与恢复生长相关性较强的相对电导率、丙二醛、游离脯氨酸、可溶性糖含量和半致死温度等5个抗寒指标进行综合评价,根据主成分的负荷量确定各个指标对抗寒性作用的权重,结合隶属度函数建立综合评价体系。最后评判出7个黄梁木种源抗寒能力强弱依次为云浮、龙州、天河、兴宁、景洪、保山、德宏,与恢复生长的试验结果基本一致。抗寒能力总体呈现广东、广西种源较云南种源强。根据前期造林对比试验,并结合本研究低温胁迫试验,在广东省新兴县、肇庆市和广州市等最低温度为-1~1 ℃的地区营造黄梁木人工林,应采用广东和广西的种源,而在广东雷州和海南海口和三亚等最低温度在1 ℃以上的地区造林,则可采用云南景洪、保山、德宏等地的种源。
目前,国内外测定抗寒性的主要方法有全株冰冻测试法(WPFT),电解质渗出率法(EL),电阻抗图谱法(EIS),叶绿素荧光法(CF)等,选择何种测定方法获得植物不同阶段抗寒性的准确信息,取决于植物组织类型和生理状况、研究目的以及已有的仪器设备[45]。在评价黄梁木抗寒能力时,应考虑对抗寒性影响的不同理化指标,利用综合评价体系进行评价,最后才能得到不同黄梁木种源抗寒能力强弱的合理结论。
| [1] |
ENDO K, OSHIMA Y, KIKUCHI H, et al. Hypotensive principles of Uncaria hooks[J]. Planta Medica, 1983, 49(3): 188-190. (  0) 0) |
| [2] |
徐英宝, 郑永光. 广东城市林业优良树种及栽培技术[M]. 广州: 广东科技出版社, 2005.
( 0) |
| [3] |
陈耀武, 管康林, 肖耀文. 团花种子休眠和萌发生理的初步研究[J]. 植物生理学通讯, 1981(6): 28-31. ( 0) |
| [4] |
任盘宇, 邹寿青. 热带速生树种团花的造林技术[J]. 林业实用技术, 2004(6): 6-8. ( 0) |
| [5] |
杨德军, 邱琼, 王达明, 等. 团花育苗技术研究[J]. 广西林业科学, 2004, 33(2): 93-95. DOI:10.3969/j.issn.1006-1126.2004.02.014 ( 0) |
| [6] |
耿云芬, 邱琼, 杨德军. 团花容器苗的育苗期施肥试验[J]. 西部林业科学, 2010, 39(1): 73-76. DOI:10.3969/j.issn.1672-8246.2010.01.014 ( 0) |
| [7] |
黄浩. 黄梁木地理种源变异与再生体系的建立[D]. 广州: 华南农业大学, 2014.
( 0) |
| [8] |
邹寿青. 团花北移试种情况调查报告[J]. 热带植物研究, 1977(11): 26-35. ( 0) |
| [9] |
许明. 团花引种试种初报[J]. 福建热作科技, 1979(4): 13-14. ( 0) |
| [10] |
郭本森. 西双版纳团花绢螟的初步研究[J]. 林业科学, 1982, 18(1): 103-106. ( 0) |
| [11] |
黄雅志, 裴汝康. 咖啡旋皮天牛和幼龄团花树[J]. 云南热作科技, 1982(2): 41-44. ( 0) |
| [12] |
韦宏. 团花树皮的吲哚生物碱成分[J]. 广西科学院学报, 1999, 15(2): 52-55. DOI:10.3969/j.issn.1002-7378.1999.02.002 ( 0) |
| [13] |
张雪. 锈毛千斤拔根及团花树皮化学成分研究[D]. 勐腊: 中国科学院西双版纳热带植物园, 2008.
( 0) |
| [14] |
钟纪育, 王文端. 团花树皮的化学成分[J]. 云南植物研究, 1990, 12(4): 453-456. ( 0) |
| [15] |
林碧珍, 张树河, 林加耕. 团花树组培快繁技术研究[J]. 中国热带农业, 2009(3): 46-47. DOI:10.3969/j.issn.1673-0658.2009.03.018 ( 0) |
| [16] |
詹艳玲. 黄梁木(Neolamarckia cadamba (Roxb. ) Bosser)组织培养研究[D]. 广州: 华南农业大学, 2010.
( 0) |
| [17] |
李娜. 团花树木葡聚糖转葡糖苷酶cDNA克隆及序列分析[D]. 北京: 北京林业大学, 2008.
( 0) |
| [18] |
马圣俊. 团花树形成层XET基因的克隆及功能初步分析[D]. 北京: 北京林业大学, 2010.
( 0) |
| [19] |
欧阳昆唏. 黄梁木α扩展蛋白的基因克隆及功能研究[D]. 北京: 北京林业大学, 2013.
( 0) |
| [20] |
云南省林科所. 滇产团花木材性质和用途的研究[J]. 云南林业科技通讯, 1975(1): 1-4. ( 0) |
| [21] |
王晶英. 植物生理生化实验技术与原理[M]. 哈尔滨: 东北林业大学出版社, 2003, 200.
( 0) |
| [22] |
李合生. 植物生理生化实验原理和技术[M]. 北京: 高等教育出版社, 2000, 279.
( 0) |
| [23] |
吕跃东, 董凤祥, 王贵禧, 等. 平欧杂交榛抗寒性综合评价体系的建立与应用[J]. 林业科学, 2008, 44(9): 31-35. DOI:10.3321/j.issn:1001-7488.2008.09.006 ( 0) |
| [24] |
何小勇. 翅荚木种源遗传多样性及其抗低温胁迫能力研究[D]. 长沙: 中南林业科技大学, 2007.
( 0) |
| [25] |
柳新红, 何小勇, 苏冬梅, 等. 翅荚木种源抗寒性综合评价体系的构建与应用[J]. 林业科学, 2007, 43(10): 45-50. ( 0) |
| [26] |
李勃, 刘成连, 杨瑞红, 等. 樱桃砧木抗寒性鉴定[J]. 果树学报, 2006, 23(2): 196-199. ( 0) |
| [27] |
朱根海, 刘祖祺, 朱培仁. 应用Logistic方程确定植物组织低温半致死温度的研究[J]. 南京农业大学学报, 1986, 9(3): 11-16. ( 0) |
| [28] |
盖钧镒. 试验统计方法[M]. 北京: 中国农业出版社, 2000.
( 0) |
| [29] |
宋锋惠, 史彦江, 吴正保, 等. 枣树枝条的抗寒性测定分析[J]. 新疆农业科学, 2009, 46(6): 1212-1215. ( 0) |
| [30] |
缴丽莉, 路丙社, 白志英, 等. 四种园林树木抗寒性的比较分析[J]. 园艺学报, 2006, 33(3): 667-670. DOI:10.3321/j.issn:0513-353X.2006.03.046 ( 0) |
| [31] |
RAJIASHEKAR C. Membrerance strutral transition: Probable relation to frost damage in hardy herbaceous species[M]. New York: Academic Press, 1979, 255-274.
( 0) |
| [32] |
戴国俊, 王金玉, 杨建生, 等. 应用统计软件SPSS拟合生长曲线方程[J]. 畜牧与兽医, 2006, 38(9): 28-30. DOI:10.3969/j.issn.0529-5130.2006.09.009 ( 0) |
| [33] |
董江水. 应用SPSS软件拟合Logistic曲线研究[J]. 金陵科技学院学报, 2007, 23(1): 21-24. ( 0) |
| [34] |
李斌, 刘立强, 罗淑萍, 等. 扁桃花芽的抗寒性测定与综合评价[J]. 经济林研究, 2012, 30(3): 16-21. ( 0) |
| [35] |
许桂芳, 张朝阳, 向佐湘. 利用隶属函数法对4种珍珠菜属植物的抗寒性综合评价[J]. 西北林学院学报, 2009, 24(3): 24-26. ( 0) |
| [36] |
郑元, 杨途熙, 魏安智, 等. 低温胁迫对仁用杏几个抗寒生理指标的影响[J]. 西北农林科技大学学报(自然科学版), 2008, 36(1): 163-167. DOI:10.3321/j.issn:1671-9387.2008.01.029 ( 0) |
| [37] |
柳新红, 何小勇, 袁德义. 中国翅荚木[M]. 北京: 中国林业出版社, 2009, 140.
( 0) |
| [38] |
陈晓阳, 沈熙环. 林木育种学[M]. 北京: 高等教育出版社, 2005, 238.
( 0) |
| [39] |
周广生, 梅方竹, 周竹青, 等. 小麦不同品种耐湿性生理指标综合评价及其预测[J]. 中国农业科学, 2003, 36(11): 1378-1382. DOI:10.3321/j.issn:0578-1752.2003.11.026 ( 0) |
| [40] |
吴行昶. 葡萄种质资源的抗寒性研究[D]. 杨凌: 西北农林科技大学, 2011.
( 0) |
| [41] |
曹建东. 9个葡萄砧木和品种的抗寒性及耐盐性鉴定[D]. 兰州: 甘肃农业大学, 2010.
( 0) |
| [42] |
张昕欣. 蜡梅品种抗寒性研究[D]. 南京: 南京林业大学, 2008.
( 0) |
| [43] |
相昆, 张美勇, 徐颖, 等. 不同核桃品种耐寒特性综合评价[J]. 应用生态学报, 2011, 22(9): 2325-2330. ( 0) |
| [44] |
孙红梅. 早实核桃对低温和干旱胁迫的生理响应及抗逆性综合评价[D]. 杨凌: 西北农林科技大学, 2012.
( 0) |
| [45] |
张钢. 国外木本植物抗寒性测定方法综述[J]. 世界林业研究, 2005, 18(5): 14-20. DOI:10.3969/j.issn.1001-4241.2005.05.004 ( 0) |