 2016, Vol. 37
2016, Vol. 37



2. 延边州水产技术推广站,吉林 延边 133002
2. The Aquaculture Technology Extending Stations of Yanbian State, Yanbian 133002, China
谷氨酰胺(Gln)是动物体内含量最丰富的氨基酸,有改善机体代谢、氮平衡、促进蛋白质合成、增加淋巴细胞数量、改善机体免疫情况以及保持肠道功能,增强机体应激反应能力的作用。因此,被人们称为“免疫营养素”[1]。有研究表明,日粮中添加一定量的Gln可以改善水产动物的生长性能、肠道功能及免疫能力[2-4]。杨奇慧等[5]对杂交罗非鱼Oreochromis niloticus♀×O.aureus♂研究表明,日粮中添加Gln可促进罗非鱼生长,提高罗非鱼抗病力,最适宜添加水平为0.6%~0.8%。Gln因溶解度低和热不稳定性,加热易生成有毒的氨和焦谷氨酸,对动物产生毒害作用,但Gln合成的谷氨酰胺二肽可以去除谷氨酰胺单体所具有的缺点,同时又具有谷氨酰胺许多的生理作用。所以,丙氨酰-谷氨酰胺(Ala-Gln)已成为生理学、营养学等学科的研究热点。有研究表明,日粮中添加Ala-Gln可以提高哲罗鱼Hucho taimen仔鱼的生长性能和抗氧化能力,且最适宜的Ala-Gln添加水平为0.75%[6]。芦洪梅等[7]研究Ala-Gln在不同密度下对建鲤Cyprinus carpio var.jian生长和饲料利用的影响,得到在20 g·L-1的养殖密度下Ala-Gln最适添加水平为0.6%。从以上试验得出,对哲罗鱼和建鲤生长、免疫的Ala-Gln的适宜添加范围为0.6%~0.75%。
γ-氨基丁酸(γ-aminobutyric acid,GABA)又被称4-氨基丁酸、γ-氨酪酸,广泛存在于自然界中,是一种非蛋白质氨基酸,在动物体内起到神经镇定作用[8-9]。具有改善大脑血液循环、降低血压、调节食欲、抵抗惊厥、抗衰老、改善肝脏、肾脏功能及对大脑缺氧和缺血有保护作用等广泛的药理作用,是一种重要的抑制性保护物质[10]。GABA作为一种安全、新型、高效的饲料添加剂,在适宜条件下对改善动物生产性能方面有着积极作用。研究表明,对动物大脑不同的区域注射一定剂量的GABA,可以显著改善动物的摄食,并且对注射剂量有着依赖效果[11]。韦习会等[12]在育肥猪饲料中添加10和20 mg·kg-1的GABA可以显著提高猪的采食量和日增质量。李庆凯等[13]对泌乳母猪研究表明,75 mg·kg-1GABA可通过刺激与采食相关的神经肽Y和胰岛素分泌提高泌乳母猪的采食量;125 g·kg-1GABA可提高泌乳母猪抗氧化能力,缓解热应激负面影响效果较好。陈黎龙等[14]对崇仁麻鸡研究表明,日粮中添加GABA可有效地改善崇仁麻鸡生长性能,且30 mg·kg-1 GABA效果最佳。戴四发[15]在10日龄的肉鸡饲料中添加Ala-Gln和GABA研究表明,饲料中添加Ala-Gln和GABA能提高试验动物采食量和日增质量,进而提高动物的生产性能。王桂芹[16]通过研究Ala-Gln对不同密度下建鲤生长和饲料利用的影响,得到在20 g·L-1的养殖密度下Ala-Gln最适添加量为6 g·kg-1。陈秀梅[17]通过研究GABA对建鲤生长性能、饲料利用及肌肉营养成分的影响,得到GABA对建鲤生长最适添加量为90 mg·kg-1。本试验选取建鲤作为研究对象,探讨在饲料中Ala-Gln和GABA最适添加量的基础上等比例增减Ala-Gln和GABA对建鲤生长、饲料利用及体成分的影响,为Ala-Gln和GABA在水产上的开发应用提供更多的理论依据。
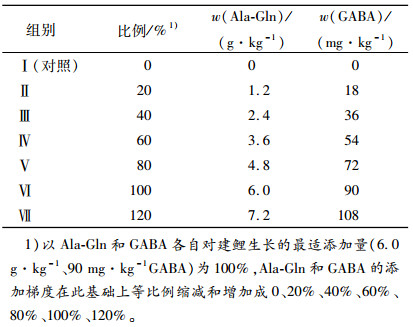
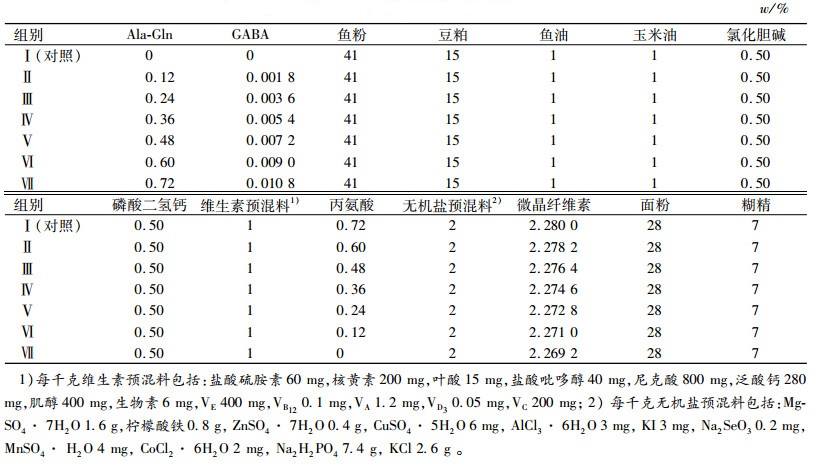
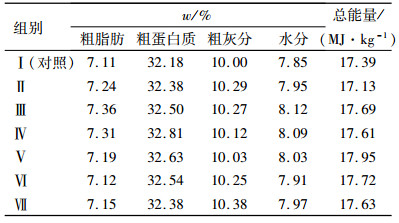
1 材料与方法 1.1 材料GABA(质量分数为99%)购于浙江省益万生物技术有限公司。Ala-Gln(质量分数为99%)购于北京莱瑞森医药科技有限公司。以鱼粉为蛋白源,配制成等氮等能(质量分数为32 %粗蛋白质、17 MJ·kg-1)饲料,在此基础上添加不同梯度Ala-Gln和GABA,共配制7组试验饲料。以Ala-Gln和GABA各自对建鲤生长的最适添加量(6.0 g·kg-1、90 mg·kg-1GABA)为100%[16-17],Ala-Gln和GABA的添加梯度是在此基础上等比例缩减和增加成0、20%、40%、60%、80%、100%、120%,具体试验设计见表 1。饲料原料经粉碎过60目筛,按配方称质量、采用逐级扩大的方法均匀混合,加工成直径为1.5 mm的颗粒,晾干后置于-20 ℃冰柜中保存、备用。采用GB/T 6435—2006、GB/T6432—1994、GB/T6438—2007和GB/T6433-2006法分别测定饲料中的水分、粗蛋白质、粗灰分和粗脂肪,弹式热量计测定能量,饲料配方见表 2,营养组成见表 3。
|
|
表 1 试验设计 Table 1 Experiment design |
|
|
表 2 试验饲料配方(干质量) Table 2 Formulation of experimental feed (dry mass) |
|
|
表 3 试验饲料营养组成(干质量) Table 3 Nutritional composition of experimental feed (dry mass) |
试验建鲤购买于吉林省东辽安西渔场。试验前建鲤暂养于吉林农业大学室内玻璃水族箱(50 cm×60 cm×80 cm)。暂养期间投喂蛋白质质量分数为32%的基础日粮作为驯化饲料,每天饱食投喂,驯化15 d,试验开始前,停止投喂1 d。
选取体质健康、大小一致(平均体质量为45.59 g)的建鲤,随机分配到21个玻璃水族箱中,共7个处理组,每个处理有3个重复,每个重复20尾。每天饱食投喂2次(09:00, 16:00),每次投喂前观察建鲤活动,投喂1 h后观察残饵情况,采用虹吸法吸出残饵,调整投喂量,记录水温、投喂量。试验期间水温为23~30 ℃,pH为7.1,溶解氧大于5.0 mg·L-1,氨氮小于0.5 mg·L-1。饲养时间为8周。
1.3 样品收集及指标计算饲养8周试验结束后,取样前停止投喂24 h,将建鲤吸干体表的水分后,每个处理组每个重复分别称质量,测体长,计算增质量率、特定生长率、饲料摄食率、饲料效率、蛋白效率及肥满度;取侧线以上、背鳍以下的白肌,测定其营养成分。
计算公式:
增质量率=(mt-m0)/m0×100%,
特定生长率=(lnmt-ln m0)/t×100%,
饲料效率=(mt-m0)/m料×100%,
摄食率=m料/[(m0+mt)/2t]×100%,
蛋白质效率=(mt-m0)/(m料×wP),
肥满度=m/L3,
其中,m0和mt分别为试验开始和试验结束时鱼体的总质量(g),t为试验的时间(d),m料为摄入饲料的干质量(g),wP为饲料蛋白质质量分数(%),m和L分别为每尾鱼的体质量(g)和体长(cm)。
1.4 统计分析采用SPSS 20.0软件进行单因素方差分析,用Duncan’s多重比较进行组间差异显著性比较,差异显著水平为P<0.05。数据用平均值±标准误表示。
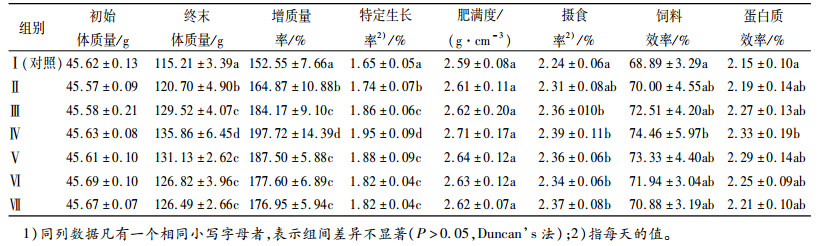
2 结果与分析 2.1 Ala-Gln和GABA对建鲤生长的影响试验Ⅱ(1.2 g·kg-1Ala-Gln、18 mg·kg-1GABA)、Ⅲ(2.4 g·kg-1Ala-Gln、36 mg·kg-1GABA)、Ⅳ(3.6 g·kg-1Ala-Gln、54 mg·kg-1GABA)组,建鲤增质量率和特定生长率随着Ala-Gln和GABA添加量的增加显著提高(P<0.05),但Ⅴ(4.8 g·kg-1Ala-Gln、72 mg·kg-1GABA)、Ⅵ(6.0 g·kg-1Ala-Gln、90 mg·kg-1GABA)、Ⅶ(7.2 g·kg-1Ala-Gln、108 mg·kg-1GABA)组建鲤平均增质量率和特定生长率不再提高,且各组之间差异不显著(表 4)。
|
|
表 4 Ala-Gln和GABA对建鲤生长及饲料利用的影响1) Table 4 Effects of Ala-Gln and GABA on growth and feed utilization of Cyprinus carpio var.jian |
将饲料中Ala-Gln和GABA添加量的和(X)与建鲤平均增质量率(Y)做抛物线回归分析,所得方程为:
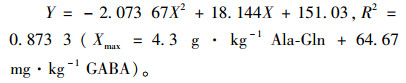
|
抛物线回归分析结果与方差分析结合起来得出,建鲤最大增质量率Ala-Gln的添加范围为3.6~4.3 g·kg-1、GABA的添加范围为54~64.67 mg·kg-1。
建鲤的形态学指标肥满度各组之间差异不显著(P>0.05)(表 4)。建鲤摄食率随着饲料中Ala-Gln和GABA添加量增加而提高,试验Ⅲ(2.4 g·kg-1Ala-Gln、36 mg·kg-1GABA)、Ⅳ(3.6 g·kg-1Ala-Gln、54 mg·kg-1GABA)、Ⅴ(4.8 g·kg-1Ala-Gln、72 mg·kg-1GABA)、Ⅵ(6.0 g·kg-1Ala-Gln、90 mg·kg-1GABA)、Ⅶ(7.2 g·kg-1Ala-Gln、108 mg·kg-1GABA)组均显著高于对照组(P<0.05),但各组之间差异不显著(P>0.05)。试验Ⅱ(1.2 g·kg-1Ala-Gln、18 mg·kg-1GABA)、Ⅲ(2.4 g·kg-1Ala-Gln、36 mg·kg-1GABA)、Ⅳ(3.6 g·kg-1Ala-Gln、54 mg·kg-1GABA)组建鲤的饲料效率和蛋白质效率都随着Ala-Gln和GABA添加量增加而提高,且第Ⅳ(3.6 g·kg-1Ala-Gln、54 mg·kg-1GABA)组建鲤蛋白质效率和饲料效率均显著高于对照组(P<0.05),Ⅴ(4.8 g·kg-1Ala-Gln、72 mg·kg-1GABA)、Ⅵ(6.0 g·kg-1Ala-Gln、90 mg·kg-1GABA)、Ⅶ(7.2 g·kg-1Ala-Gln、108 mg·kg-1GABA)组,饲料效率和蛋白质效率均不再提高且各组之间差异不显著(P>0.05)(表 4)。
将饲料中Ala-Gln和GABA添加量的和(X)与建鲤蛋白质效率(Y)做抛物线回归分析,所得方程为:
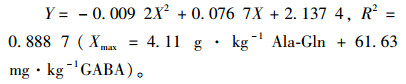
|
抛物线回归分析结果与方差分析结合起来得出,建鲤最大饲料效率的最适Ala-Gln添加范围为3.6~4.11 g·kg-1、GABA的添加范围为54~61.63 mg·kg-1。
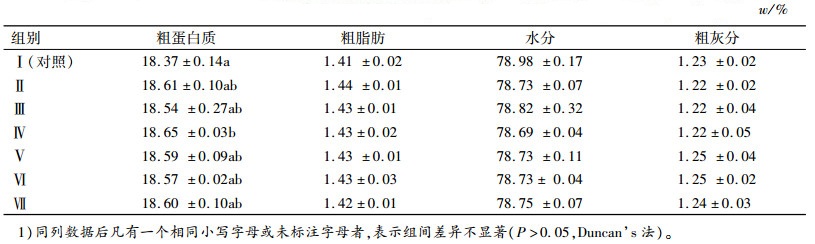
2.2 Ala-Gln和GABA对建鲤肌肉营养成分的影响试验Ⅱ(1.2 g·kg-1Ala-Gln、18 mg·kg-1GABA)、Ⅲ(2.4 g·kg-1Ala-Gln、36 mg·kg-1GABA)、Ⅳ(3.6 g·kg-1Ala-Gln、54 mg·kg-1GABA)、Ⅴ(4.8 g·kg-1Ala-Gln、72 mg·kg-1GABA)、Ⅵ(6.0 g·kg-1Ala-Gln、90 mg·kg-1GABA)、Ⅶ(7.2 g·kg-1Ala-Gln、108 mg·kg-1GABA)组建鲤肌肉中粗蛋白质含量随着饲料中Ala-Gln和GABA添加量的增加呈上升趋势,且第Ⅳ组蛋白质含量显著高于对照组(P<0.05),但Ⅱ~Ⅶ各组之间差异不显著(P>0.05)。建鲤肌肉中的粗灰分、粗脂肪、水分各组之间差异不显著(P>0.05)(表 5)。
|
|
表 5 Ala-Gln和GABA对建鲤肌肉营养成分的影响 Table 5 Effects of Ala-Gln and GABA on nutrition composition in the muscle of Cyprinus carpio var.jian |
本试验中,所有添加Ala-Gln和GABA的组建鲤生长均显著高于对照组,说明外源性添加Ala-Gln和GABA能促进建鲤生长。Ala-Gln能有效维持肠道正常形态,不断地为水产动物肠黏膜细胞的更新提供能量和代谢前体,且提高其消化酶的活性,从而改善机体消化机能,减少因外界不利因素造成的水产动物生产性能下降,促进水产动物的生长[18]。朱青等[19]在德国镜鲤C.carpio日粮中添加0.25%~1.5%、0.75%Ala-Gln能显著提高德国镜鲤鱼体粗蛋白质含量、增质量率、蛋白质效率、饲料效率和蛋白质沉积率(P<0.05),表明日粮中添加Ala-Gln能提高鱼体蛋白质的沉积和生长。GABA不仅是参与味觉调制的神经递质,而且还参与了调节动物采食行为和味觉感受,从而促进水产动物食欲,提高其采食量。曹德瑞等[20]在肥育猪日粮中添加GABA,以10 mg·kg-1 GABA效果最为显著,料质量比降低,提高日增质量、眼肌面积和屠宰率,不同剂量GABA对育肥猪内脏器官未产生显著影响。陈强等[21]在断奶仔猪日粮中添加GABA,结果显示15 mg·kg-1 GABA能提高断奶仔猪平均日采食量和平均日增质量。在本试验中,Ala-Gln和GABA添加组建鲤摄食率、蛋白质效率和饲料效率随着Ala-Gln和GABA添加量增加而提高,说明饲料中添加Ala-Gln和GABA能提高饲料和蛋白质的利用率,从而促进生长。
本试验中,3.6 g·kg-1Ala-Gln、54 mg·kg-1GABA添加组和4.8 g·kg-1Ala-Gln、72 mg·kg-1GABA添加组比6.0 g·kg-1Ala-Gln、90 mg·kg-1GABA添加组(即所添加的Ala-Gln和GABA均为对建鲤的最适添加量)效果显著。从体内物质生化代谢关系分析,Ala-Gln和GABA均属于氨基酸类物质,它们之间存在着密切的代谢相关性。Ala-Gln在体内被肠黏膜消化吸收有2种途径:①Ala-Gln在细胞外经胞膜二肽酶水解释放出Gln,再被肠黏膜消化吸收;②Ala-Gln被转运到上皮细胞内,经胞质内的二肽酶水解释放出Gln。Gln和GABA在一定的代谢条件下可以相互转化,并与TAC建立联系。Gln在谷氨酰胺酶作用下,生成谷氨酸(Glutamate,Glu)。而谷氨酸在谷氨酸脱羧酶的作用下,生成GABA。所以,Gln是GABA合成前体,在细胞培养液中加入Gln可以促进GABA的形成,当GABA不足时,Gln被更多地利用。从生理功能方面分析,Gln主要从营养、供能、解毒和抗氧化等方面为机体提高抗应激能力。GABA则主要从抑制神经兴奋、降低血压和抗惊厥等神经调控方面提高机体的抗应激能力,显示了二者在提高动物生产性能和抗应激功能方面的共性和调控方式的互补性。这些都与本试验结果一致,Ala-Gln和GABA促进动物生长作用,可能是通过促进动物生长相关激素的合成与释放,进而调节机体代谢,具体机理有待进一步的研究。
3.2 Ala-Gln和GABA对建鲤肌肉营养成分的影响本研究中随着Ala-Gln和GABA添加量增加,建鲤肌肉中粗蛋白质含量先呈上升后呈下降的趋势,且3.6 g·kg-1Ala-Gln、54 mg·kg-1GABA添加组显著高于对照组。建鲤肌肉中的粗脂肪、水分、粗灰分含量各组之间差异不显著。
由此来看,饲料中添加Ala-Gln和GABA有促进肌肉中蛋白质合成作用,这可能与Ala-Gln和GABA调节机体内分泌水平有关。GABA是一种抑制性神经递质,通过下丘脑-垂体-性腺轴系来影响性腺和垂体的生理机能,从而参与激素的分泌调节[22]。研究表明,GABA具有促进生长激素(GH)分泌的作用,而GH具有可促进动物生长、提高机体内蛋白质沉积、降低脂肪生成的作用[23],GABA对内分泌功能尤其对腺垂体激素分泌功能有着非常重要的调节作用,口服5 g GABA可以使血液中生长激素显著提高[24]。Fan等[25]研究发现,GABA不仅能有效改善猪的生产性能, 还能显著促进与动物生长、摄食及代谢相关的激素(生长激素、促甲状腺激素、褪黑素)分泌。Gln是机体内组织间氨基转化作用的重要载体;Gln是核酸、核苷、蛋白质、氨基酸等生物分子合成的前体;Gln是蛋白质代谢的重要调节因子, 能促进细胞内蛋白质的合成, 减少骨骼肌中蛋白质的分解[26];Gln可以通过对内分泌调控促进养殖动物生长。在动物机体内,Gln可转化为精氨酸(Arg),Arg浓度增加刺激GH分泌,GH提高DNA聚合酶活性,促进mRNA转录,从而加速了蛋白的表达[27]。朱青等[19]在镜鲤日粮中添加Ala-Gln,结果发现添加Ala-Gln试验组镜鲤血清中总蛋白(TP)含量高于不添加Ala-Gln的对照组,这表明Ala-Gln可以促进镜鲤幼鱼体内蛋白质的合成。全鱼营养成分分析显示,添加Ala-Gln试验组镜鲤鱼体蛋白质含量均高于不添加Ala-Gln的对照组,进一步证明Ala-Gln可提高镜鲤的蛋白质合成。Gln通过细胞增容作用,促进肌细胞的生长和分化,刺激体内生长激素、胰岛素的分泌,使机体的合成代谢大于分解代谢。Gln可以改变某些激素如类胰岛素生长因子(IGF-I)和甲状腺素含量。谢建新等[28]对大鼠、黄冠庆等[29]对断奶仔猪研究表明,Gln可以提高机体内游离三碘甲腺原氨酸(FT3)和游离甲状腺素(FT4)含量,还可以通过上调小肠黏膜细胞IGF-I mRNA表达量,增加IGF-I自分泌和旁分泌,促进蛋白质合成,从而促进生长。
在本试验条件下,饲料中添加Ala-Gln和GABA在一定程度上促进了建鲤幼鱼的生长,并对促进肌肉中蛋白质的合成发挥了一定的作用,其中以Ala-Gln添加量为3.6 g·kg-1、GABA添加量为54 mg·kg-1的添加组效果最佳。以增质量率和饲料效率作为评价指标,经过回归方程预测,建鲤配合饲料中最适Ala-Gln添加范围为3.6~4.11 g·kg-1、GABA添加范围为54~64.63 mg·kg-1。
| [1] |
MELIS G C, WENQE N, BOELENS P G, et al. Glutamine:Recent developments in research on the clinical significance of glutamine[J]. Curr Opin Clin Nutr Metab Care, 2004, 7(1): 59-70. DOI:10.1097/00075197-200401000-00011 (  0) 0) |
| [2] |
叶元土, 王永玲, 蔡春芳, 等. 谷氨酰胺对草鱼肠道L-亮氨酸、L-脯氨酸吸收及肠道蛋白质合成的影响[J]. 动物营养学报, 2007, 19(1): 28-32. DOI:10.3969/j.issn.1006-267X.2007.01.005 ( 0) |
| [3] |
姜俊, 周小秋. 谷氨酰胺对肠上皮细胞的调节作用[J]. 饲料工业, 2004, 25(4): 31-33. ( 0) |
| [4] |
徐奇友, 王常安, 许红, 等. 外源性谷氨酰胺对虹鳟稚鱼生长和肠道形态的影响[J]. 中国粮油学报, 2009, 24(4): 98-102. ( 0) |
| [5] |
杨奇慧, 周歧存, 谭北平. 谷氨酰胺对杂交罗非鱼生长、饲料利用及抗病力的影响[J]. 中国水产科学, 2008, 15(6): 1017-1023. ( 0) |
| [6] |
徐奇友, 王常安, 许红, 等. 饲喂丙氨酰-谷氨酰胺对哲罗鱼仔鱼生长和抗氧化能力的影响[J]. 动物营养学报, 2009, 21(6): 1012-1017. DOI:10.3969/j.issn.1006-267x.2009.06.032 ( 0) |
| [7] |
芦洪梅, 韩宇田, 王桂芹, 等. 丙胺酰-谷氨酰胺对不同养殖密度建鲤生长和饲料利用的影响[J]. 华南农业大学学报, 2013, 34(2): 241-247. ( 0) |
| [8] |
LEE B J, KIM J S, KANG Y M, et al. Antioxidant activity and γ-aminobutyric acid (GABA) content in sea tangle fermented by Lactobacillus brevis BJ20 isolated from traditional fermented foods[J]. Food Chem, 2010, 122(1): 271-276. DOI:10.1016/j.foodchem.2010.02.071 ( 0) |
| [9] |
KARDOS J. Recent advances in GABA research[J]. Neurochemi Internati, 1999, 34(5): 353-358. DOI:10.1016/S0197-0186(99)00036-4 ( 0) |
| [10] |
郑红发, 黄亚辉, 刘霞林, 等. γ-氨基丁酸的药理作用[J]. 茶叶通讯, 2004(4): 14-18. DOI:10.3969/j.issn.1007-1253.2004.04.011 ( 0) |
| [11] |
SHUYE P U, MUKUL R J, TAMAS L, et al. Interactions between neuropepitde Y and γ-aminobutyric acid in stimulation of feeding: A morphological and pharmacological analysis[J]. Endocrinology, 1999, 140(2): 933-940. DOI:10.1210/endo.140.2.6495 ( 0) |
| [12] |
韦习会, 漆兴桂, 夏东, 等. 日粮添加GABA对育肥猪生长和饲料利用的影响[J]. 家畜生态, 2004, 25(2): 11-13. ( 0) |
| [13] |
李庆凯, 李同洲, 李宁宁, 等. γ-氨基丁酸对泌乳母猪采食量和抗氧化能力的试验[J]. 单胃动物营养, 2010(3): 49-52. ( 0) |
| [14] |
陈黎龙, 游金明, 黄建珍, 等. γ-氨基丁酸对崇仁麻鸡生长性能和血液生理生化指标的影响[J]. 动物营养学报, 2010, 22(2): 452-459. DOI:10.3969/j.issn.1006-267x.2010.02.031 ( 0) |
| [15] |
戴四发. 谷氨酰胺和γ-氨基丁酸对肉鸡抗应激和肉品质的影响及机理探讨[D]. 南京: 南京农业大学, 2012.
( 0) |
| [16] |
王桂芹. 谷氨酰胺对拥挤胁迫鲤鱼的营养调控及抗胁迫机理的研究[D]. 长春: 吉林农业大学, 2010.
( 0) |
| [17] |
陈秀梅. γ-氨基丁酸对建鲤生长、免疫和抗氨氮胁迫的影响[D]. 长春: 吉林农业大学, 2015.
( 0) |
| [18] |
孙浪, 刘臻, 周玲, 等. 谷氨酰胺二肽在水产动物营养中的应用研究[J]. 饲料广角, 2012(5): 45-47. DOI:10.3969/j.issn.1002-8358.2012.05.031 ( 0) |
| [19] |
朱青, 徐奇友, 王长安, 等. 丙氨酰-谷氨酰胺对德国镜鲤幼鱼(Cyprinus carpio L.)血清生化指标及体组成的影响[J]. 水产学杂志, 2009, 22(4): 12-15. DOI:10.3969/j.issn.1005-3832.2009.04.003 ( 0) |
| [20] |
曹德瑞, 邹晓庭. γ-氨基丁酸对生长肥育猪生长性能和胴体品质的影响[J]. 饲料工业, 2007, 28(23): 31-34. DOI:10.3969/j.issn.1001-991X.2007.23.011 ( 0) |
| [21] |
陈强, 梁军生. γ-氨基丁酸对断奶仔猪生产性能及血清蛋白水平和酶活性的影响研究[J]. 中国农学通报, 2012, 28(35): 47-50. DOI:10.3969/j.issn.1000-6850.2012.35.010 ( 0) |
| [22] |
包华琼, 王新庄. 氨基丁酸(GABA)的生殖生理作用[J]. 动物医学进展, 2002, 23(3): 39-40. DOI:10.3969/j.issn.1007-5038.2002.03.012 ( 0) |
| [23] |
王涛. γ-氨基丁酸饲用价值研究进展[J]. 中国畜牧杂志, 2010, 46(6): 61-63. ( 0) |
| [24] |
MCCANN S M, VIJAYAN E, NEGRO-VILAR A, et al. GAMMA aminobutyric acid (GABA), a modulator of anterior pituiary hormone secretion by hypothalamic and pituitary action[J]. Psychoneuroendocrinology, 1984, 9(2): 97-106. DOI:10.1016/0306-4530(84)90029-5 ( 0) |
| [25] |
FAN Z Y, DENG J P, LIU G H, et al. Study on the effect of γ-aminobutyric acid on the reproduction performance and internalh ormone levels in growing pigs[J]. Chin J Anim Nutr, 2007, 19(4): 350-356. ( 0) |
| [26] |
袁玖, 姚军虎. 谷氨酰胺的生物学功能及其在断奶仔猪中的应用[J]. 饲料工业, 2005, 26(8): 8-11. DOI:10.3969/j.issn.1001-991X.2005.08.003 ( 0) |
| [27] |
周光明, 王嫒, 吴西华, 等. 谷氨酰胺的生物学功能研究进展及其在养牛生产中的应用[J]. 中国奶牛, 2013(1): 13-16. DOI:10.3969/j.issn.1004-4264.2013.01.004 ( 0) |
| [28] |
谢建新, 顾岩, 刘银坤, 等. 联合应用生长激素和谷氨酰胺对短肠大鼠小肠黏膜吸收功能的影响[J]. 解剖学杂志, 2001, 24(3): 231-234. ( 0) |
| [29] |
黄冠庆, 黄晓亮, 李嘉嘉, 等. 丙氨酰谷氨酰胺对断奶仔猪生长和血清抗氧化能力的影响[J]. 动物营养学报, 2008, 20(6): 706-711. DOI:10.3969/j.issn.1006-267X.2008.06.016 ( 0) |