 2016, Vol. 37
2016, Vol. 37



东京黄檀Dalbergia tonkinensis俗称越南黄花梨、越南香枝木、通京黄檀等[1-2],为蝶形花科Papilionaceae黄檀属Dalbergia中的一种速生半落叶乔木,一般生长在海拔400~800 m的天然林及次生林中。原产地越南(集中在河内、永福、兴安、凉山、北河、广平和宁平等省)、泰国及老挝等地通常叫做Mai Dou Lai,意为“有花纹的花梨木”。根据越南林业部的标准2198-CNR《全国木材通用分类表》,越南名为“sue”的木材即为越南黄花梨,并于2006年3月将其正式列为IA组保护树种,对其进行挂牌标识保护。经中国林业科学研究院专家分析鉴定,将其木材暂归为我国《红木》标准(GB/T 18107—2000)的香枝木类[3]。作为天然林濒临灭绝的另一种香枝木,与降香黄檀D. odorifera存在诸多共性,其木材坚硬珍贵,纹理清晰美观,多鬼眼,同具辛辣酸味,并且生长快速,成为降香黄檀理想的替代品而倍受关注。
国内外有关东京黄檀树种的研究报道并不多,我国只是从2002起相继从泰国、越南、柬埔寨等国家初步引进东京黄檀的种子进行引种驯化和栽培等基础性研究。陈美红等[4]在育苗技术方面做了相关工作,刘小金等[5]研究了温度对东京黄檀种子发芽率的影响,并指出其最适温度为30 ℃左右,吴师强等[2]就越南黄花梨叶、花、果等方面进行了初步的表征观察研究,鲜见其他相关的引种或栽培技术的系统研究报道。本文以降香黄檀为对照,对从越南引进的东京黄檀10个产地47个家系的种子,开展了种子形态变异及播种品质差异性研究,以揭示东京黄檀种子形态在产地和家系间的差异以及种子生理特性的不同,旨在为其选择优良种质及进一步的育种和遗传改良等基础性研究提供参考。
1 材料与方法 1.1 试验材料试验材料包括从越南Gia Lai、Hưng Yên、Lạng Sơn、Bắc Ninh、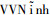
|
|
表 1 试验种子产地及基本情况 Table 1 Basic information of different seed provenances |
将各家系的种子随机选取100粒,平均分成4份,用电子游标卡尺分粒测量种子纵轴为种长(L)、横向最宽处为种宽(W)、种子背腹两面最宽处为种厚(H),并计算种子长宽比(L: W)和厚宽比(H: W),进行描述统计及方差分析。
1.2.2 种子品质指标测定根据《林木种子检验规程GB2772—99》种子品质指标的测定要求[6],本试验测定千粒质量(百粒法,3次重复)、发芽率、发芽势和发芽指数等相关指标。
1.2.3 种子发芽试验试验于2014年6月份进行。将已随机均分的各家系种子均匀播种于已编号的15 cm×25 cm育苗盘中,播种前苗床器皿、种子先用蒸馏水洗净,用质量分数为0.5%的高锰酸钾喷施进行表面杀菌10 min,再用蒸馏水洗净后播种于河沙基质中(太阳暴晒后喷施质量分数为0.5%的高锰酸钾)。2 d后每隔24 h观测记录种子发芽情况,定期喷水保持基质相当的湿度。连续2 d没有种子发芽时结束统计。发芽时间(d)从置床播种之日起计算,发芽势计算至发芽高峰日为止。发芽指标的计算公式如下[6]:

|
原始数据用Excel 2007输入整理,用SPSS 20.0的ANOVA对种子形态指标和发芽特征进行显著性检验,用相关分析(Correlate)中的Person系数法分析种子形态特征、千粒质量、发芽特征及地理环境因子等的相关关系,用系统聚类法(Hierarchical cluster)对种子产地进行初步区划,并用Origin 8.0作图。
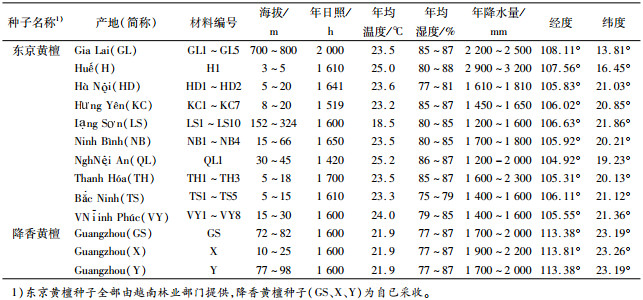
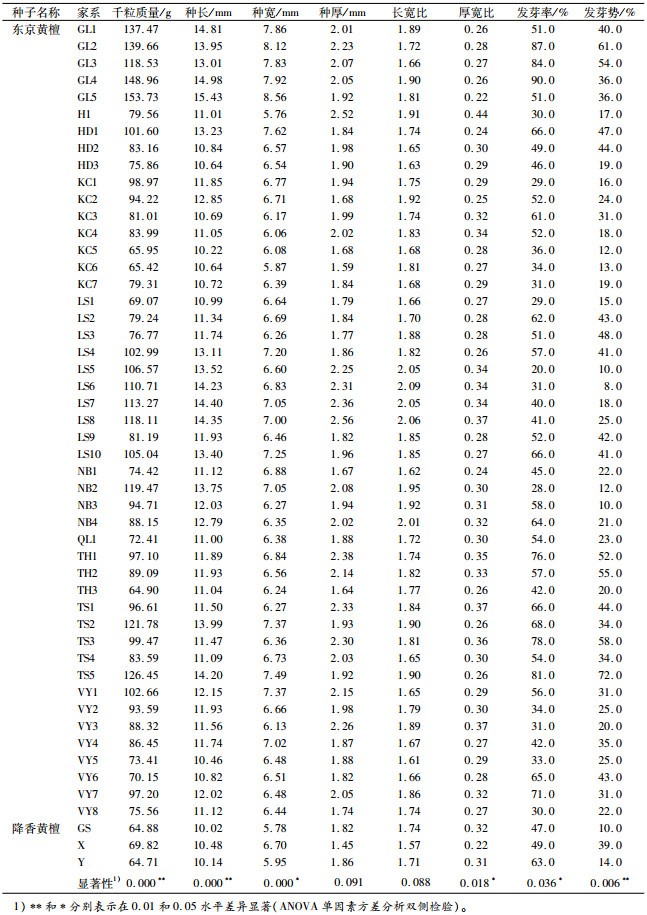
2 结果与分析 2.1 不同产地/家系东京黄檀种子形态差异不同产地/家系种子形态和发芽特征测定结果见表 2。东京黄檀的种子长10.22~15.43 mm,宽5.76 ~8.56 mm,厚1.59~2.56 mm;对比降香黄檀种子长10.02~10.48 mm,宽5.78~6.70 mm,厚1.45~1.86 mm,相对变幅较大,并有长、宽、厚均大于降香黄檀的趋势。东京黄檀种长均值最大的是GL产地(14.26 mm)(表 3),其中尤为突出的是GL5(15.43 mm)和GL4(14.98 mm)家系,最小的为QL产地(10.98 mm),相对增长30%;种子宽最大值GL产地(8.03 mm)是最小值H产地(5.76 mm)的1.39倍;而种厚最大的种子是H产地(2.52 mm),最小的是KC产地(1.83 mm),相对增厚38%。长宽比和厚宽比最大的是H产地。种长变异系数最大的产地是HD、LS和TS(0.12),最小的为QL和TH(0.07);种宽和种厚变异系数最大的产地分别为TS(0.10)和LS(0.18), 最小的分别为QL(0.05)、和H(0.09),且除H产地外均大于或等于降香黄檀。不同产地的种宽变异系数都低于或是等于相应的种长和种厚的变异系数,且以种厚变异系数最大。
|
|
表 2 不同产地家系的黄檀种子形态与发芽特征 Table 2 The seed morphology and germination characteristics of different provenances and families of Dalbergia |
|
|
表 3 不同产地黄檀种子形态特征1) Table 3 Seed morphology of different provenances of Dalbergia |
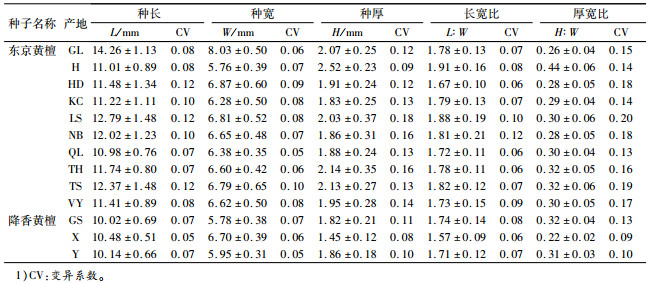
各家系的均值比较结果显示:GL产地的5个家系的种子长、宽、厚都大于其他产地各家系(表 2),其变异范围也较小。其他产区各家系种子长基本在12.00 mm左右,种宽、种厚除了个别之外,分别在6.50 mm和1.80 mm左右。
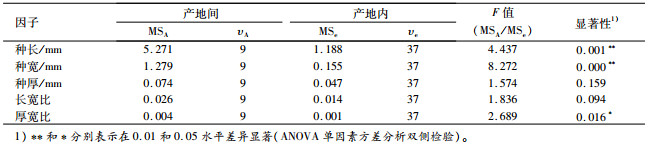
东京黄檀产地间的种子形态表型性状方差分析结果(表 4)显示,种子长和宽达到极显著差异(P<0.01),种子厚和长宽比差异不显著(P>0.05),厚宽比差异显著(P<0.05)。这与前人对西伯利亚红松Pinus sylvestris、白皮松Pinus bungeana、厚朴Magnolia officinalis、白桦Betula platyphylla、小桐子Jatropha curcas、大叶栎Castanopsis fissas等树种不同种源及家系种子性状的变异研究结果基本一致[7-13]。产地间极显著的种子长宽大小变异,表明东京黄檀产地间的种子大小形态的表型稳定性存在着丰富的性状遗传变异[14]。
|
|
表 4 不同产地东京黄檀种子形态特征方差分析 Table 4 Analysis of variance on seed morphology of different provenances of Dalbergia tonkinensis |
种长、种宽和种厚在不同产地之间存在着一定的趋势关系,这很可能是在遗传和环境的双向选择下形成的一种变化规律[15]。不同产地种子大小测定结果表明,不管是东京黄檀还是降香黄檀,种长、种宽都存在相似的变化趋势,即种宽随着种长的消涨而消涨,表现为显著的正相关关系(图 1);种厚与种长、种宽的变化规律则不尽相同,不因种长而厚,不因种宽而厚,甚至出现相反的结果(图 1)。

|
图 1 不同产地黄檀种子种长、种宽和种厚的关系 Figure 1 The relationship among seed length, width and thickness of different provenances of Dalbergia |
种子含水量一致时的千粒质量是衡量种子品质最重要的指标,千粒质量可以反映种子饱满度,并且间接反映种子大小、营养物质的含量等[16]。这与种子发芽、苗木建植都有直接或间接的相关关系[17]。发芽指数则是一项与发芽率有相似变化规律的综合指标,在发芽时间和发芽数这两个维度上能较充分地阐释种子的萌发情况。试验结果(表 5)表明,东京黄檀不同产地之间其种子千粒质量存在极显著差异(P<0.01),发芽率、发芽势和发芽指数存在显著或极显著差异。
|
|
表 5 不同产地黄檀种子发芽特征及千粒质量差异比较 Table 5 Comparisons of seed germination characteristics and 1 000-seed mass of different provenances of Dalbergia |
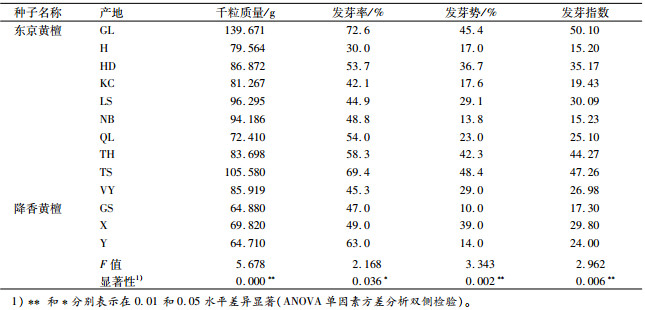
千粒质量最大的是GL产地,达到139.671 g,其发芽率(72.6%)和发芽指数(50.10)也都大于其他产地的种子,并且其各家系的千粒质量、发芽率和发芽势多数大于其他产区。其次为TS产地,干粒质量为105.580 g,同样对应较高的发芽率(69.4%)、发芽势(48.4%)和发芽指数(47.26),其各家系的发芽特征也仅次于GL产区部分家系。千粒质量最小的产地为QL,干粒质量为72.410 g,然而其发芽率等却并非最低,这可能与种子饱满度密切相关,试验发现当种子长宽厚比例相当(表 3、图 1),种子较圆满紧实时,种子能储存较丰富的营养物质而利于发芽率的提高[13, 15, 17]。东京黄檀种长、种宽最大的GL产地、较大的TS产地及种长最小的QL产地,其长宽比、厚宽比均相当(表 3),表现为种子饱满,其相对应的就有较高的发芽率。
从种子形态特征和千粒质量与发芽情况的相关关系结果(表 6)可见,发芽率与千粒质量及种子宽度极显著正相关(P<0.01),与种子长度显著正相关(P<0.05),而种子厚度对发芽率、发芽势和发芽指数的贡献相对较小(P>0.05)。这也说明种子大小与种子发芽情况有着极为密切的关系,相当的种子大小比例是影响种子发芽建植至关重要的因素[18]。
|
|
表 6 东京黄檀种子形态特征、发芽特征、千粒质量与地理、气候因子的相关关系1) Table 6 Correlations among seed morphology, seed germination characteristics, 1 000-seed mass of Dalbergia tonkinensis and geographic and climatic factors |
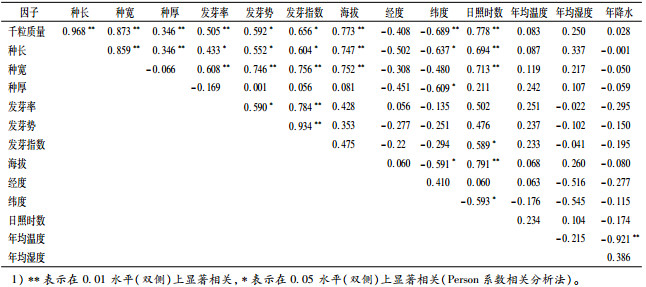
进一步分析发现东京黄檀种子的千粒质量、种长和种宽与海拔和日照时数呈现极显著正相关,而与纬度分别呈极显著、显著和不显著负相关,与经度不显著负相关;而种子发芽特征与产地的地理和环境因子相关性基本不显著。
2.4 东京黄檀原产地种子的聚类分析种子产地的区划对引种至关重要[10],本试验中东京黄檀各产地间种子特征差异显著。由此以种子形态(种长、种宽、种厚)、千粒质量、发芽特征(发芽率、发芽势、发芽指数)等为因子进行产地的类平均法(组内联结法)的平方欧氏距离聚类分析。结果如图 2所示,东京黄檀不同产地可以简单地划分为3个区:GL产区; 以LS和TS产地为代表,并包括TH、VY及HD产地的大区;以KC和NB产地为代表,包括QL和H产地的大区。根据种子形态特征及发芽试验结果,如果从种子质量和播种品质角度出发选择引种产地,可初步优先选择GL产区的种子,其次为TS产区的种子。

|
图 2 基于种子生物学特征的东京黄檀类平均法聚类分析 Figure 2 Hierarchical cluster analysis of different Dalbergia tonkinensis families based on seed biological characteristics |
种子是植物生活史较为稳定和基础的特征之一,种子的大小(长、宽、厚)与产地有关[8, 13],更与后期种子传播、休眠、发芽及幼苗建成等密切相关[19-21],并且大小种子各有利弊[15]。所以根据种子大小及萌发情况对引种产地进行选择,成为外引树种必不可少的基础性工作。
对引进的东京黄檀10个产地的种子试验研究表明:不同产地东京黄檀的种长、种宽、长宽比、厚宽比、千粒质量以及发芽特征都达到了极显著或显著的水平,这说明对其引种产地很有必要进行选择。试验结果表明,产自GL产地的种子优势突出,它们有最大的千粒质量、种长和种宽、发芽率和发芽指数,其次为TS产地。种子的饱满度则在一定程度上影响着播种品质。
种子发芽是反应种子生活力强弱的重要因素之一[22],且较直观易测。对东京黄檀及降香黄檀不同产地种子的发芽试验表明,其发芽率、发芽势、发芽指数等都达到显著性差异水平,这种发芽差异性可为选择适应性优良的种子进行引种提供参考。
试验表明,千粒质量、种子长、种子宽与发芽率、发芽势都显著正相关,其中种宽与发芽率、发芽势、发芽指数相关性都达到极显著水平,根据这些种子表型性状与种子发芽的关系,可以为前期引种东京黄檀并培育优质苗木提供一个良好的理论支持。
环境条件对种子形态特征的可塑性同样具有非常重要的作用,有研究指出种子大小、千粒质量等与经纬度之间有着显著相关性[10-12],本试验中,种子形态特征(种长、种宽及千粒质量)与纬度显著负相关,与经度不显著负相关。这与刘桂丰等[10]对白桦种子研究得出的结论一致,而与海拔和日照时数呈现极显著正相关,这说明东京黄檀种子受海拔、日照时数长的影响较为明显。通过对引进的10个产地的东京黄檀种子形态及发芽试验研究,发现GL产地的种子在各方面表现均较为理想,但其后续的表现则仍需开展进一步的苗期试验及造林生长观测方能给以定论。
| [1] |
中国科学院中国植物志编辑委员会. 中国植物志:第40卷[M]. 北京: 科学出版社, 1994, 121.
(  0) 0) |
| [2] |
吴师强, 李大周, 陆文, 等. 越南黄花梨形态特性及引种植物名称研究初报[J]. 热带林业, 2013, 41(3): 25-26. DOI:10.3969/j.issn.1672-0938.2013.03.009 ( 0) |
| [3] |
国家质量技术监督局. 红木: GB/T18107—2000[S]. 北京: 中国标准出版社, 2000.
( 0) |
| [4] |
陈美红, 刘小金, 徐大平, 等. 越南黄花梨育苗技术[J]. 林业实用技术, 2014(7): 33-34. ( 0) |
| [5] |
刘小金, 徐大平, 杨曾奖, 等. 温度对越南黄花梨种子萌发的影响[J]. 林业科学研究, 2014, 27(5): 707-709. ( 0) |
| [6] |
国家质量技术监督局. 林木种子检验规程: GB2772—1999[S]. 北京: 中国标准出版社, 1999: 116.
( 0) |
| [7] |
王成. 西伯利亚红松引种与种源试验研究[D]. 哈尔滨: 东北林业大学, 2011. http://cdmd.cnki.com.cn/Article/CDMD-10225-1011146728.htm
( 0) |
| [8] |
李晓洁, 徐化成. 白皮松种子发芽习性及其种源变异的研究[J]. 林业科学, 1989, 25(2): 97-105. DOI:10.3321/j.issn:1001-7488.1989.02.012 ( 0) |
| [9] |
杨志玲, 杨旭, 谭梓峰, 等. 厚朴不同种源及家系种子性状的变异[J]. 中南林业科技大学学报, 2009, 29(5): 49-55. ( 0) |
| [10] |
刘桂丰, 孔红娃. 白桦不同种源种子形态特征及发芽率[J]. 东北林业大学学报, 1999, 27(4): 1-4. DOI:10.3969/j.issn.1000-5382.1999.04.001 ( 0) |
| [11] |
栗宏林, 张志翔, 张鑫. 小桐子不同产地种子性状及苗期生长差异研究[J]. 干旱区资源与环境, 2010, 24(2): 204-208. ( 0) |
| [12] |
李春, 樊东函, 梁机, 等. 大叶栎种子性状与苗期生长地理种源变异[J]. 广西科学, 2009, 16(2): 210-214. ( 0) |
| [13] |
舒枭, 杨志玲, 杨旭, 等. 不同产地厚朴种子性状的变异分析[J]. 林业科学研究, 2010, 23(3): 457-461. ( 0) |
| [14] |
程诗明, 顾万春. 苦楝表型性状梯度变异的研究[J]. 林业科学, 2006, 42(5): 29-35. ( 0) |
| [15] |
武高林, 杜国祯. 植物种子大小与幼苗生长策略研究进展[J]. 应用生态学报, 2008, 19(1): 191-197. ( 0) |
| [16] |
黄双全, 刘桂霞, 韩建国. 种子大小和播种深度对种苗建植的影响[J]. 草业科学, 2007, 24(6): 44-49. DOI:10.3969/j.issn.1001-0629.2007.06.011 ( 0) |
| [17] |
武高林, 杜国祯, 尚占环. 种子大小及其命运对植被更新贡献研究进展[J]. 应用生态学报, 2006, 17(10): 1969-1972. DOI:10.3321/j.issn:1001-9332.2006.10.037 ( 0) |
| [18] |
宗文杰, 刘坤, 卜海燕, 等. 高寒草甸51种菊科植物种子大小变异及其对种子萌发的影响研究[J]. 兰州大学学报(自然科学版), 2006, 42(5): 52-55. DOI:10.3321/j.issn:0455-2059.2006.05.011 ( 0) |
| [19] |
HARPER J L, LOVELL P H, MOORE K G. The shapes and sizes of seeds[J]. Annu Rev Ecol Evol Syst, 1970, 1(1): 327-356. DOI:10.1146/annurev.es.01.110170.001551 ( 0) |
| [20] |
FEMMER M. Relationships between seed weight ash content and seedling growth in twenty-four species of Compositae[J]. New Phytol, 1983, 95(4): 697-706. DOI:10.1111/nph.1983.95.issue-4 ( 0) |
| [21] |
SILVERTOWN J. Seed size, life span, and germination date as co-adapted features of plant life history[J]. Am Nat, 1981, 118(6): 860-864. DOI:10.1086/283876 ( 0) |
| [22] |
朱念德. 植物学:形态解剖结构[M]. 广州: 中山大学出版社, 2000, 179-180.
( 0) |