 2016, Vol. 37
2016, Vol. 37



2. 安徽农业大学 理学院,安徽 合肥 230036
2. School of Science, Anhui Agricultural University, Hefei 230036, China
卵形短须螨Brevipalpus obovatus寄主为茶树Camellia sinensis L.、山茶树C.japonica L.、咖啡树Coffea arabica 等45科120种植物, 主要为害老叶和成叶叶背, 使叶片失去光泽, 主脉褐变, 叶背多紫褐色突起小斑, 后期叶柄霉变脱落, 削弱茶树生长势[1]。王宏毅[2]在福建南部发现卵形短须螨为害西番莲Passiflora caerulea L., 并分析了降雨和温度对其田间种群数量消长的影响。朱梅等[3]对广东地区茶场的卵形短须螨进行了研究, 释放捕食螨优势种江原钝绥螨Amblyseius eharai对卵形短须螨的防治效果达23.30%~81.31%。双斑长跗萤叶甲Monolepta hieroglyphica成虫、幼虫均可危害茶树, 是近年南方茶区为害成灾的新害虫[1, 4]。王立仁等[5]通过研究为害玉米等作物的双斑长跗萤叶甲, 提出了抢种、深翻和施药相结合的防治措施。李广伟等[6]研究了温度对双斑长跗萤叶甲成虫寿命及生殖力的影响。梁日霞等[7]对来自中国北方26个不同地理种群的双斑长跗萤叶甲线粒体细胞色素C氧化酶亚基Ⅱ(COⅡ)基因片段序列核苷酸多态性的研究表明, 这些地区双斑长跗萤叶甲的不同地理种群间已发生明显的遗传分化。
这2种害虫的防治多采用化学和物理方法, 化学防治中的滥用农药现象, 不仅导致茶叶农药残留超标, 害虫也已经产生抗药性[8]。保护和利用天敌是持续控制茶树害虫的重要措施[9]。Sinu[10]研究了印度北部茶园中以鸟类为主的混合群落对茶园害虫的抑制作用。Chen等[11]研究表明在茶园中间作不同植被能有效增加蜘蛛的数量, 从而抑制害虫的发生。本文研究白毫早茶树C.sinensis cv.Baihaozao和乌牛早茶树C.sinensis cv.Wuniuzao园2种主要害虫卵形短须螨和双斑长跗萤叶甲与其天敌的关系, 以期为其生物防治提供科学依据。
1 材料与方法 1.1 材料调查地点为安徽农业大学农业科技示范园茶园(31°94′N, 117°21′E), 茶树品种为白毫早和乌牛早, 调查的茶园面积均为0.2 hm2。调查时间为2015年3月28日至11月14日, 每半个月调查1次, 共调查17次。
白毫早茶树由湖南省农业科学院茶叶研究所从安化群体中单株育种而成, 茶树芽叶生育力强、生长速度快、产量高, 春茶萌发期在3月上旬, 适宜在长江南北绿茶茶区栽植。
乌牛早茶树中的永嘉乌牛早茶是近年的新创名茶, 特早发芽, 3月上旬即可采制, 用它加工的乌牛早龙井, 持嫩性较强, 适宜浙江省尤其是扁形类名优茶产区作早生搭配品种推广[12-13]。
1.2 方法 1.2.1 取样方法采用平行跳跃法, 在茶园随机选取3行, 每行间隔1 m取1个2 m长的样方, 每行10个样方, 用沾有洗衣粉水液的搪瓷盘(40 cm×30 cm)对样方中所有枝条进行盘拍, 每样方随机选取10片叶片, 共300片叶片, 调查并记录害虫及其天敌物种数和个体数。
1.2.2 2种害虫与其天敌在数量关系上的灰色关联度分析[14]将2种害虫数量(Yi)及其主要天敌数量(Xj)分别看作一个本征性系统, 2种害虫数量(Y1)和(Y2)作为该系统的参照序列。对2种害虫(Y1)和(Y2)与天敌(Xj)在不同时点上第k点的效果白化值, 进行双序列关系分析, 经数据均值化后得:
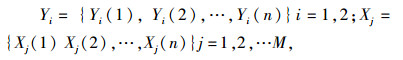
|
Yi与Xj在第k点的关联系数rij(k)为:
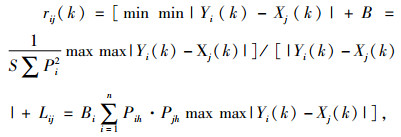
|
式中, k=1, 2, …, n, ρ为分辨系数, 取值区间[0, 1], 一般取ρ=0.5, 为扩大各关联度之间的差异, 本文取ρ=0.8。R(Yi, Xj)=1/n∑rij(k)为第j种天敌(Xj)与卵形短须螨(Y1)和双斑长跗萤叶甲(Y2)的关联度, 其大小反映Xj对Yi的联系或影响程度[15]。
1.2.3 天敌与害虫的时间及空间生态位分析用Levins[16]的生态位重叠指数公式和Morisita相似性系数公式[13]计算天敌与害虫在空间和时间上生态位重叠指数和相似性系数, 来判断天敌对害虫在空间和时间上跟随关系的密切程度。
Levins的生态位宽度指数公式:
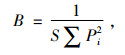
|
式中, B为物种的生态位宽度; Pi 为物种利用第i等级资源占利用总资源的比例; S为资源系列的等级数。
Levins的生态位重叠指数公式:

|
式中, Lij为物种i对物种j的生态位重叠, Pih和Pjh为每个物种在资源序列的第h单位上的比例, Bi为物种i的生态位宽度。
生态位相似性比例采用Morisita相似性系数公式[17]:
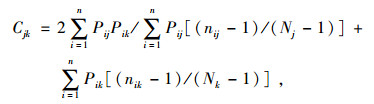
|
式中, Pij, Pik 分别表示种j, k在第i个资源等级上所占的比例, nik是k物种在i资源序列等级上的数量, nij 是j物种在i资源序列等级上的数量。Nj , Nk分别表示j物种和k物种的个体数量之和。
1.2.4 天敌与其目标害虫关系的综合分析将所得的天敌与目标害虫数量、时间及空间的关联参数分别除以本项参数的最大值, 其商为密切指数, 密切指数相加, 和最大即为目标害虫的第1位天敌, 依次类推, 最后对2种茶园2种主要害虫的前6位天敌进行比较[11, 18]。
2 结果与分析 2.1 茶园卵形短须螨和双斑长跗萤叶甲及其天敌种群数量动态2个品种茶园的卵形短须螨和双斑长跗萤叶甲及数量较多的9种捕食性天敌和2种寄生性天敌种群消长动态见表 1。卵形短须螨2015年有2个发生高峰, 4月11日—5月10日是第1个高峰期, 10月11日—11月14日是第2个高峰期。双斑长跗萤叶甲2015年有2个发生高峰, 5月10日—6月20日是第1个高峰期, 10月11日—11月14日是第2个高峰期。天敌中鳞纹肖蛸Tetragnatha squamata、锥腹肖蛸Tetragnatha maxillosa、八点球腹蛛Theridion octomaculatum、粽管巢蛛Oxyopes sertatus和日本追寄蝇Exorita japonica较多。
|
|
表 1 白毫早和乌牛早茶园卵形短须螨和双斑长跗萤叶甲及其天敌种群动态1) Table 1 Dynamics of the numbers of Monolepta hieroglyphica, Brevipalpus obovatus and their natural enemies in Baihaozao and Wuniuzao tea gardens |
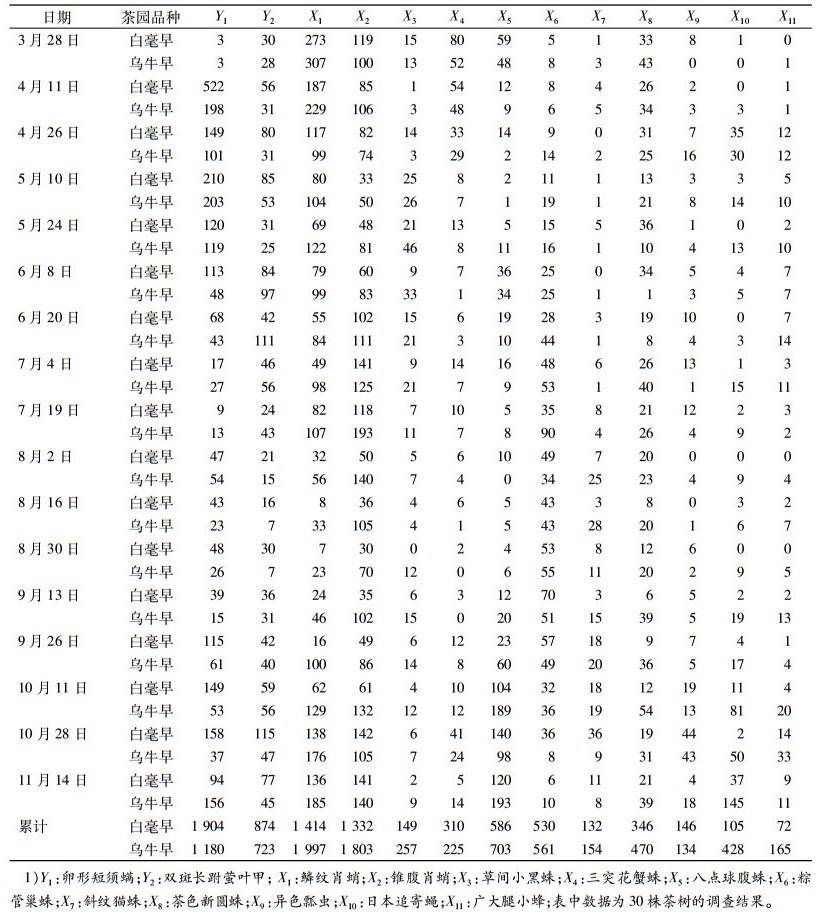
对2个茶园品种害虫和天敌的种群数量差异进行t检验, 结果为:ν=32, t0.05=2.04, t0.01=2.75, 卵形短须螨(1.283 8);双斑长跗萤叶甲(0.929 2);鳞纹肖蛸(1.402 2);锥腹肖蛸(2.153 0);草间小黑蛛Erigonidium graminicolum(1.953 5);三突花蟹蛛Misumenops tricuspidatus(0.769 3);八点球腹蛛(0.372 8);粽管巢蛛(0.246 7);斜纹猫蛛Clubiona japonicola(0.414 8);茶色新圆蛛Neoscona theisi(1.790 2);异色瓢虫Harmonia axyridis (0.197 4);日本追寄蝇(2.026 8);广大腿小蜂Brachymeria obscurata(2.515 4)。可以看出2个品种茶园2种害虫之间差异均不显著, 11种天敌中只有锥腹肖蛸(2.153 0)和广大腿小蜂(2.515 4)大于t0.05, 差异显著。
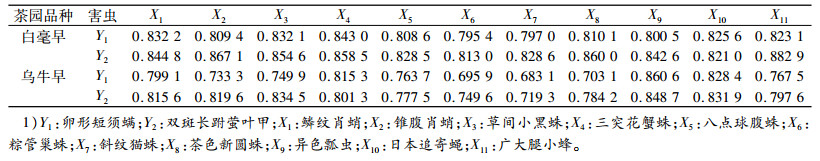
2.2 白毫早和乌牛早茶园卵形短须螨和双斑长跗萤叶甲与其天敌在数量上的关系对茶园中卵形短须螨和双斑长跗萤叶甲与其天敌在数量关系上的关联度分析结果(表 2)表明, 白毫早茶园中与卵形短须螨关联度最大的前6位天敌依次是三突花蟹蛛、鳞纹肖蛸、草间小黑蛛、日本追寄蝇、广大腿小蜂和茶色新圆蛛; 双斑长跗萤叶甲的是广大腿小蜂、锥腹肖蛸、茶色新圆蛛、三突花蟹蛛、草间小黑蛛和鳞纹肖蛸。乌牛早茶园中与卵形短须螨关联度最大的前6位天敌依次是异色瓢虫、日本追寄蝇、三突花蟹蛛、鳞纹肖蛸、广大腿小蜂和八点球腹蛛; 与双斑长跗萤叶甲关联度最大的前6位天敌依次是异色瓢虫、草间小黑蛛、日本追寄蝇、锥腹肖蛸、鳞纹肖蛸和三突花蟹蛛。
|
|
表 2 白毫早和乌牛早茶园卵形短须螨和双斑长跗萤叶甲与其主要天敌的关联度1) Table 2 Correlation degrees between Monolepta hieroglyphica and Brevipalpus obovatus and their natural enemies in Baihaozao and Wuniuzao tea gardens |
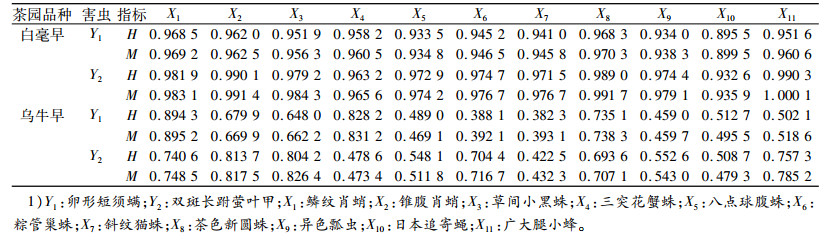
卵形短须螨和双斑长跗萤叶甲与其天敌时间生态位重叠指数(H)和相似性比例(M)结果见表 3。总时间资源单位为17个。白毫早茶园与卵形短须螨生态位重叠指数最大的天敌是鳞纹肖蛸(0.968 5), 相似性比例最大的是茶色新圆蛛(0.970 3);与双斑长跗萤叶甲的生态位重叠指数和相似性比例最大的均是广大腿小蜂(0.990 3和1.000 1)。乌牛早茶园卵形短须螨的天敌均是鳞纹肖蛸(0.894 3和0.895 2);与双斑长跗莹叶甲生态位重叠指数最高的是锥腹肖蛸(0.813 7), 相似性比例最大的是草间小黑蛛(0.826 4)。
|
|
表 3 卵形短须螨和双斑长跗萤叶甲与其天敌时间上的Horn’s生态位重叠指数(H)和Morisita相似性比例(M) 1) Table 3 The time niche overlap indices represented as Horn's indices (H) and proportional similarities represented as Morisita indices (M) of Monolepta hieroglyphica and Brevipalpus obovatus and their natural enemies |
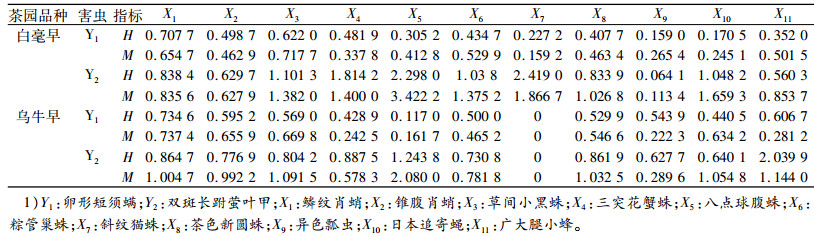
本研究以5月10日的数据计算卵形短须螨和双斑长跗萤叶甲与其天敌空间生态位重叠指数H和相似性比例M, 结果见表 4。白毫早茶园与卵形短须螨空间生态位重叠指数最大的是鳞纹肖蛸(0.707 7), 相似性比例最大的是草间小黑蛛(0.717 7);双斑长跗萤叶甲空间生态位重叠指数、相似性比例最大的均是八点球腹蛛(2.419 0和3.422 2)。乌牛早茶园与卵形短须螨的M、H最大的均是鳞纹肖蛸(0.734 6和0.737 4);与双斑长跗萤叶甲生态位重叠指数最高的是广大腿小蜂(2.039 9), 相似性比例最大的是八点球腹蛛(2.080 0)。
|
|
表 4 卵形短须螨和双斑长跗萤叶甲与其天敌的空间上的Horn’s生态位重叠指数(H)和Morisita相似性比例(M) 1) Table 4 The spatial niche overlap indices represented as Horn's indices (H) and proportional similarities represented as Morisita indices (M) of Monolepta hieroglyphica and Brevipalpus obovatus and their natural enemies |
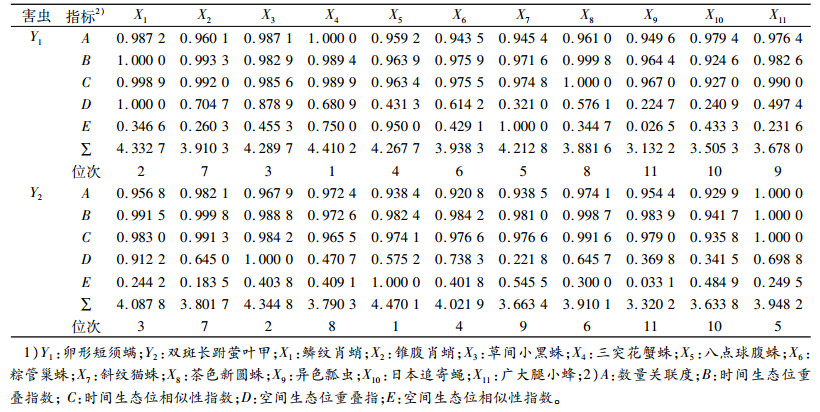
将白毫早茶园卵形短须螨和双斑长跗萤叶甲与其天敌在数量、时间、空间关系的密切指数之和列于表 5。密切指数之和越大, 表明该天敌与害虫关系越密切。卵形短须螨前6位的优势种天敌的是三突花蟹蛛、鳞纹肖蛸、草间小黑蛛、八点球腹蛛、斜纹猫蛛和粽管巢蛛; 双斑长跗萤叶甲的是八点球腹蛛、草间小黑蛛、鳞纹肖蛸、粽管巢蛛、广大腿小蜂和茶色新圆蛛。
|
|
表 5 白毫早茶园卵形短须螨和双斑长跗萤叶甲与其天敌关系的密切指数1) Table 5 Correlation indices of Monolepta hieroglyphica and Brevipalpus obovatus and their natural enemies in Baihaozao tea garden |
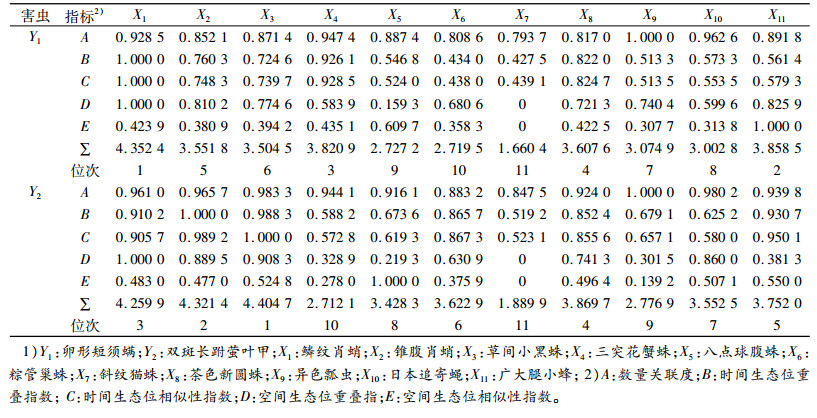
乌牛早茶园卵形短须螨和双斑长跗萤叶甲与其天敌在数量、时间、空间关系的密切指数之和见表 6。卵形短须螨前6位的优势种天敌是鳞纹肖蛸、广大腿小蜂、三突花蟹蛛、茶色新圆蛛、锥腹肖蛸和草间小黑蛛; 双斑长跗萤叶甲的是草间小黑蛛、锥腹肖蛸、鳞纹肖蛸、茶色新圆蛛、广大腿小蜂和粽管巢蛛。
|
|
表 6 乌牛早茶园卵形短须螨和双斑长跗萤叶甲与其天敌关系的密切指数1) Table 6 Correlation indices of Monolepta hieroglyphica and Brevipalpus obovatus and their natural enemies in Wuniuzao tea garden |
为比较茶树品种对害虫优势种天敌的影响, 将2个品种茶园卵形短须螨和双斑长跗萤叶甲的11种天敌中的前6位优势种天敌列于表 7。可以看出, 卵形短须螨的11种主要天敌的前6位优势种天敌的变化, 茶树品种间的累计相同比率除第1位外, 其余均维持在50%。而双斑长跗萤叶甲的11种主要天敌的前6位优势种天敌的累计相同比率为83.33%, 随着天敌种类的增加累计相同比率也增加。表明茶树品种对双斑长跗萤叶甲优势种天敌的影响比对卵形短须螨优势种天敌的影响小。
|
|
表 7 白毫早和乌牛早茶园卵形短须螨和双斑长跗萤叶甲前6位优势种天敌比较 Table 7 Comparisons of the top six dominant natural enemies of Monolepta hieroglyphica and Brevipalpus obovatus in Baihaozao and Wuniuzao tea gardens |
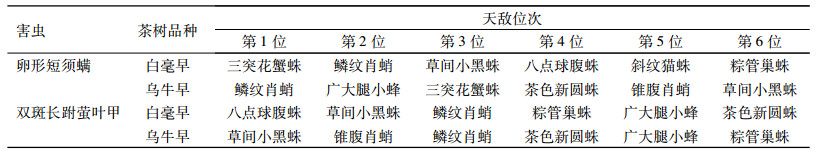
运用灰色关联度分析法、生态位分析法和综合密切指数法, 综合评判2015年白毫早和乌牛早2个品种茶园卵形短须螨和双斑长跗萤叶甲的优势种天敌, 并对茶园中害虫优势种天敌种群进行分析比较, 本研究中共有11种与卵形短须螨和双斑长跗萤叶甲有关且数量较多的天敌, 通过对2种主要害虫与11种天敌在数量、时间和空间关系上的综合分析, 卵形短须螨在白毫早和乌牛早茶园的前6位优势种天敌中, 相同的天敌是三突花蟹蛛、鳞纹肖蛸和草间小黑蛛, 相同天敌比率为50%。双斑长跗萤叶甲在白毫早和乌牛早茶园的前6位优势种天敌中, 相同的天敌是草间小黑蛛、鳞纹肖蛸、粽管巢蛛、茶色新圆蛛和广大腿小蜂, 相同天敌比率为83.33%。卵形短须螨优势种天敌的累计相同比率均维持在50%左右。而双斑长跗萤叶甲优势种天敌的累计相同比率随着天敌种类的增加而增加。表明2个茶树品种对卵形短须螨优势种天敌的影响大于对双斑长跗萤叶甲优势种天敌的影响。
影响天敌种群动态的因子包括食饵(害虫)、天敌的天敌以及无机环境因子等。这些因子以食饵作用最大, 食饵又受寄主植物影响, 抗感性不同的寄主影响害虫的种群数量和空间分布。将影响天敌种群因子的作用权重, 若寄主植物和害虫占70%, 依此为据分析茶树品种对2种害虫优势种天敌的影响, 若令茶树品种对天敌优势种没有影响, 则累计相同天敌比率应为70%, t检验法分析表明卵形短须螨的优势种天敌的t值为2.944 0, 当υ=5时, t0.05=2.57, t0.01=4.03, 可知t>t0.05, 白毫早茶园与乌牛早茶园对卵形短须螨优势种天敌的影响差异显著; 双斑长跗萤叶甲的优势种天敌的t值为1.593 3, t<t0.05, 2个茶树品种之间对双斑长跗萤叶甲优势种天敌影响的差异不显著。若把累计相同天敌比率提高至75%, 则卵形短须螨和双斑长跗萤叶甲优势种天敌的t值分别为3.484 7和2.027 8, 其结论不变。为了比较2个品种茶树对2种害虫优势种天敌影响的差异, 进一步对卵形短须螨和双斑长跗萤叶甲优势种天敌累计相同比率t检验, t=0.549 6<t0.05(2.23)。尽管茶树品种对2种害虫优势种天敌有影响, 但差异不显著。
茶树品种对害虫天敌优势种的影响, 究其原因, 可能是2个茶树品种的生物物理条件和生物化学条件不同, 导致茶树品种对昆虫抗感性的差异, 使害虫的数量和发生规律产生变化进而影响到天敌种群的数量变化, 这也符合邓欣等[19]的研究。茶园昆虫群落随时间、茶园类型、茶树空间等不同条件而变化的动态特征[20-22], 反映了天敌对害虫的跟随关系。茶树品种对害虫天敌优势种的影响机制有待进一步探讨。
| [1] |
张汉鹄, 谭济才. 中国茶树害虫及其无公害治理[M]. 合肥: 安徽科学技术出版社, 2004, 49-51.
(  0) 0) |
| [2] |
王宏毅. 卵形短须螨为害西番莲研究[J]. 福建农林大学学报(自然科学版), 2002, 31(3): 320-323. ( 0) |
| [3] |
朱梅, 侯柏华, 吴伟南, 等. 茶园螨类调查及利用胡瓜钝绥螨控制卵形短须螨的初步研究[J]. 环境昆虫学报, 2010, 32(2): 204-209. DOI:10.3969/j.issn.1674-0858.2010.02.010 ( 0) |
| [4] |
聂强, 孙强. 双斑萤叶甲成虫的取食选择性研究[J]. 黑龙江八一农垦大学学报, 2009, 21(4): 38-41. DOI:10.3969/j.issn.1002-2090.2009.04.010 ( 0) |
| [5] |
王立仁, 刘斌侠, 付泓. 玉米田双斑长跗萤叶甲的发生为害情况与防治对策[J]. 陕西农业科学, 2006(2): 123-131. DOI:10.3969/j.issn.0488-5368.2006.02.060 ( 0) |
| [6] |
李广伟, 陈秀琳, 张建萍, 等. 温度对双斑长跗萤叶甲成虫寿命及繁殖的影响[J]. 昆虫知识, 2010, 47(2): 322-325. ( 0) |
| [7] |
梁日霞, 王振营, 何康来, 等. 基于线粒体COⅡ基因序列的双斑长跗萤叶甲中国北方地理种群的遗传多样性研究[J]. 昆虫学报, 2011, 54(7): 828-837. ( 0) |
| [8] |
郑龙章, 张春霞, 黄森慰. 茶农使用农药行为影响因素实证研究:以福建省为例[J]. 福建农林大学学报(哲学社会科学版), 2009, 12(2): 44-49. DOI:10.3969/j.issn.1671-6922.2009.02.010 ( 0) |
| [9] |
邹运鼎. 害虫管理中的天敌评价理论与应用[M]. 北京: 中国林业出版社, 1997, 27-90.
( 0) |
| [10] |
SINU P A. Avian pest control in tea plantations of sub-Himalayan plains of Northeast India:Mixed-species foraging flock matters[J]. Biol Control, 2011, 58(3): 362-366. DOI:10.1016/j.biocontrol.2011.05.019 ( 0) |
| [11] |
CHEN L L, YOU M S, CHEN S B. Effects of cover crops on spider communities in tea plantations[J]. Biol Control, 2011, 59(3): 326-335. DOI:10.1016/j.biocontrol.2011.09.007 ( 0) |
| [12] |
姚国坤. 中国名优茶地图[M]. 上海: 上海文化出版社, 2013, 84-85.
( 0) |
| [13] |
徐海荣. 中国茶事大典[M]. 北京: 华夏出版社, 2000, 200-201.
( 0) |
| [14] |
刘思峰, 谢乃明. 灰色系统理论及其应用[M]. 6版. 北京: 科学出版社, 2013, 48-87.
( 0) |
| [15] |
郭骅, 周夏芝, 毕守东, 等. 茶园黑刺粉虱和茶叶甲成虫与其捕食性天敌之间的关系[J]. 华南农业大学学报, 2013, 34(3): 345-351. ( 0) |
| [16] |
FU W G, WANG F K, YIN Q L, et al. Niche dynamics of species in succession process in the wetland of Yangtze Rivers lower reach, China[J]. Plant Ecol Evol, 2015, 148(1): 43-51. DOI:10.5091/plecevo ( 0) |
| [17] |
TAKADA Y. Morisita's prosperity index revisited[J]. Biodivers Conserv, 2015, 24(8): 2093-2097. DOI:10.1007/s10531-015-0933-5 ( 0) |
| [18] |
OLLERTON J, STOTT A, ALLNUTT E, et al. Pollination niche overlap between a parasitic plant and its host[J]. Oecologia, 2007, 151(3): 473-485. DOI:10.1007/s00442-006-0605-y ( 0) |
| [19] |
邓欣, 谭济才. 生态控制茶园内害虫、天敌种类及数量的季节变化规律[J]. 生态学报, 2002, 22(7): 1166-1172. DOI:10.3321/j.issn:1000-0933.2002.07.028 ( 0) |
| [20] |
韩宝瑜. 三类典型茶园昆虫和螨类群落组成和动态的差异[J]. 茶叶科学, 2005, 25(4): 249-254. DOI:10.3969/j.issn.1000-369X.2005.04.003 ( 0) |
| [21] |
柯胜兵, 党凤花, 毕守东, 等. 不同海拔茶园害虫、天敌种群及其群落结构差异[J]. 生态学报, 2011, 31(14): 4161-4168. ( 0) |
| [22] |
陈亦根, 熊锦君, 黄明度, 等. 复合茶园昆虫类群多样性和稳定性研究[J]. 华南农业大学学报, 2004, 25(1): 59-61. DOI:10.3969/j.issn.1001-411X.2004.01.016 ( 0) |