 2016, Vol. 37
2016, Vol. 37



2. 华南农业大学 林学与风景园林学院/广东省森林植物种质资源创新与利用重点实验室, 广东 广州 510642
2. College of Forestry Science and Landscape Architecture, South China Agricultural University/ Guangdong Key Laboratory for Innovative Development and Germplasm Utilization of Forest Plant, Guangzhou 510642, China
香椿Toona sinensis又名红香椿、椿芽树、香椿头、椿甜树等, 是我国特有的速生优质用材树种[1], 主要分布于我国大陆, 在台湾地区也有分布[2]。现有的报道多集中在食用、药用方面的研究[3-4], 也有不少栽培和木材材性与利用方面的研究[5-10]。由于香椿分布广泛, 对其进行种源研究是有必要的, 但现有相关文献较少, 而对于其种源光合特性的研究更少。张玉洁[11]做了香椿幼树光合作用及其影响因子研究; 杨玉珍等[12]研究了干旱胁迫对6个不同种源香椿苗木光合特性的影响; 郝明灼等[13]做了不同种源香椿芽菜品质分析及光合特性比较; 孟丽等[14]研究了Cd2+、Hg2+污染对香椿叶绿素含量和光合特性的影响。为此, 本研究在分布区均匀布点, 采集11省14县(市、区)的香椿种子, 对各种源1年生苗木5个瞬时光合生理指标进行测定和分析, 旨在丰富香椿种源光合特性的研究, 为优良香椿种源的筛选提供依据。
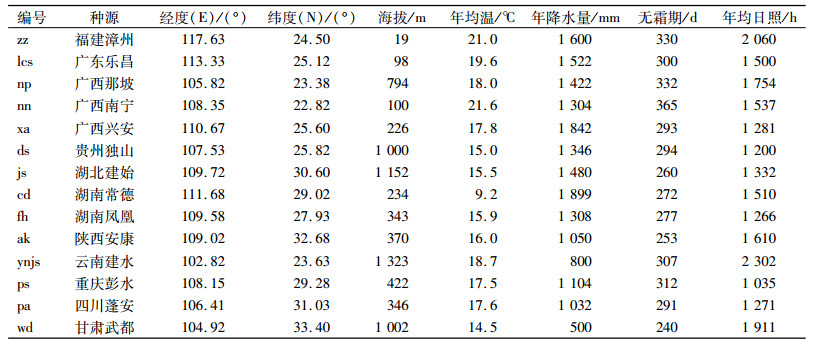
1 材料与方法 1.1 试验材料从我国福建漳州、广东乐昌、广西那坡、广西南宁、广西兴安、贵州独山、湖北建始、湖南常德、湖南凤凰、陕西安康、云南建水、重庆彭水、四川蓬安、甘肃武都11个省(市、区)14个县采集香椿种子, 香椿种子的具体情况见表 1。于2013年3月9日在华南农业大学试验地播种, 12月9日定植于苗圃。
|
|
表 1 香椿种源采种点地理位置和气候因子状况 Table 1 Geographic locations and climatic information of the seed collection sites of different Toona sinensis provenances |
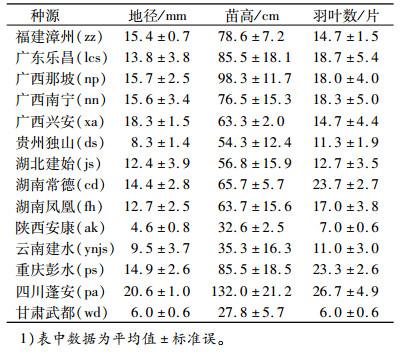
在2014年7月下旬, 选择晴朗天气, 于上午09:00~12:00用Li-6400便携式光合作用分析系统(美国LI-COR)对14个1年生香椿种源苗测定叶片瞬时净光合速率(Pn)、气孔导度(Gs)、胞间CO2浓度(Ci)、蒸腾速率(Tr)。供试各种源随机选取生长中庸、无病虫害、生长势基本一致的香椿苗3株, 每株选择中部发育正常的活体功能羽叶1~2片, 选择靠顶部的3片小叶进行测定。苗木基本生长情况见表 2。测定过程中采用人工叶室, 使用Li-6400-2B红蓝光源, 光强设置为1 200 μmol·m-2·s-1, 温度35 ℃, CO2 400 μmol·mol-1, 测定每片小叶时读取10个数据, 取平均值作为该叶片在该时间点的光合参数值, 最后对平均值进行分析。
|
|
表 2 香椿苗木生长情况1) Table 2 Growth status of Toona sinensis seedlings |
通过以下公式计算出苗木叶片瞬时水分利用效率(WUE):
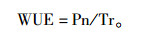
|
(1) |
数据采用Excel2007整理, Spss19.0进行方差分析、相关分析、主成分分析。方差分析采用固定模型。为了解各性状在种源间的差异和遗传稳定性, 通过下式计算变异系数(CV)和重复力(R):
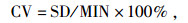
|
(2) |
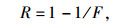
|
(3) |
式中, SD是性状标准差, MIN是性状平均数, F为方差分析中种源间与误差的方差比。
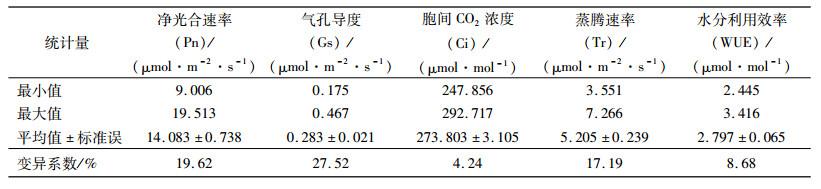
2 结果与分析 2.1 香椿种源瞬时光合生理指标变异统计香椿种源的各个瞬时光合生理指标变异结果见表 3, 各个指标在种源间存在变异。5个指标的变异系数变动幅度为4.24%~27.52%。其中, Gs变异系数最大, Gs变异幅度为0.175~0.467 μmol·m-2·s-1, 说明香椿光合生理指标在种源间的差异是明显的。而Ci的变异系数最小, 为4.24%, 变异幅度247.856~292.717 μmol·mol-1, 说明该性状在种源间的差异较小。
|
|
表 3 香椿种源主要光合生理指标变异统计 Table 3 Summary of variation in photosynthetic physiological indices of different Toona sinensis provenances |
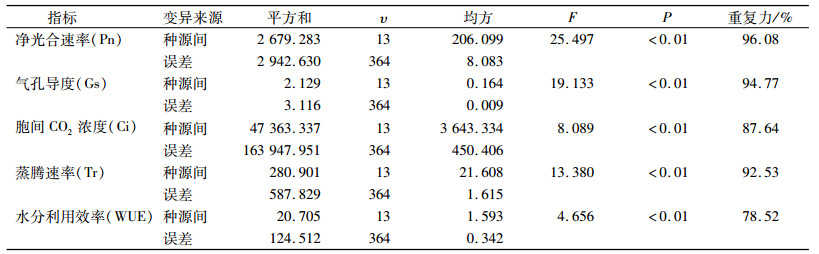
方差分析(表 4)表明, 香椿5个光合生理指标在种源间都存在极显著差异。F值大小顺序为Pn(25.497)>Gs(19.133)>Tr(13.380)>Ci(8.089)>WUE(4.656)。5个参数的重复力变动范围为78.52%~96.08%。其中, Pn重复力最高, 为96.08%, 其次是Gs、Tr, 均高达90%以上, WUE重复力最低, 但也达到78.52%。由此说明, 光合参数主要由遗传因子控制, 可作为香椿性状遗传多样性评价和地理变异研究的主要性状。
|
|
表 4 不同种源香椿光合生理指标方差分析 Table 4 Analysis of variance of photosynthetic physiological indices of different Toona sinensis provenances |
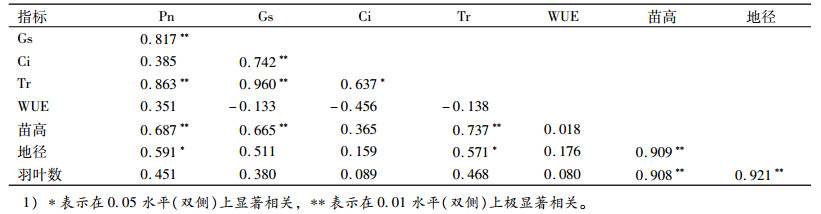
香椿种源光合生理、生长指标相关性分析结果(表 5)表明:Pn与Gs、Tr和苗高呈极显著正相关, Pn与地径呈显著正相关; Gs与Ci、Tr和苗高呈极显著正相关; Ci与Tr呈显著正相关; Tr与苗高呈极显著正相关, Tr与地径呈显著正相关; 苗高与地径、羽叶数呈极显著正相关; 地径与羽叶数呈极显著正相关。
|
|
表 5 光合生理指标和生长指标的相关系数1) Table 5 Correlation coefficient between photosynthetic physiological indices and growth indices |
由表 6可以看出:广西兴安、广西那坡、四川蓬安香椿种源的Pn较大, 分别达到了19.513、17.641、16.543 μmol·m-2·s-1, 云南建水、湖南常德、甘肃武都种源的Pn较小, 分别为9.006、10.415、11.837 μmol·m-2·s-1; Gs最大的是广西那坡、广西兴安、四川蓬安种源, 分别为0.467、0.372、0.356 μmol·m-2·s-1, 最小的是云南建水、甘肃武都、湖南常德种源, 分别为0.181、0.175、0.208 μmol·m-2·s-1; Ci值的差异相对较小, 其中陕西安康、广西那坡种源的Ci最大, 分别为292.717、291.837 μmol·mol-1, 甘肃武都种源的Ci最小, 为247.856 μmol·mol-1; Tr最大的是广西那坡、四川蓬安、广西兴安种源, 分别为7.266、6.081、6.044 μmol·m-2·s-1, 最小的是云南建水、甘肃武都种源, 分别为3.551、4.128 μmol·m-2·s-1; WUE最大的是广西兴安、广东乐昌种源, 分别为3.416、3.038 μmol·mol-1, 最小的是广西那坡种源, 为2.445 μmol·mol-1。
|
|
表 6 不同种源香椿的叶片光合生理指标差异比较1) Table 6 Comparisons of photosynthetic physiological indices of different Toona sinensis provenances |
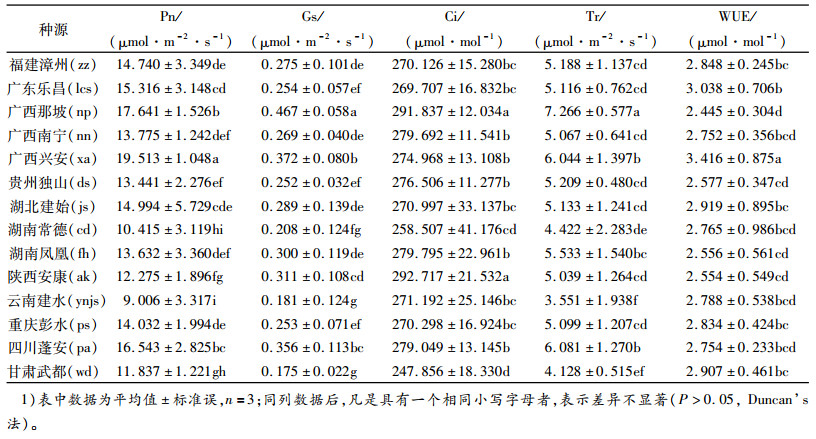
对14个香椿种源的光合生理指标进行主成分分析, 对各种源数据采用无量纲方法中的均值化[15], 选择最大方差法得到旋转后的主成分特征向量与特征根(表 7)。由表 7可知:主成分1(PRIN1)、主成分2(PRIN2)贡献率分别为63.397%、29.686%, 二者累计贡献率为93.083%;Pn、Gs、Tr、Ci是PRIN1的主要因子, 都有很大的正载荷; Ci、Tr、Gs、WUE是PRIN2的主要因子, 其中WUE、Pn为正载荷, 其他3个参数为负载荷。
|
|
表 7 香椿种源光合生理指标主成分分析的特征向量和特征根 Table 7 Characteristic vectors and characteristic roots from the principle component analysis of the photosynthetic physiological indices of Toona sinensis provenances |
对香椿各个种源的光合生理指标进行主成分转换, 其结果见图 1。可把14个种源分为3类:第1类为甘肃武都(wd)、湖南常德(cd)和云南建水(ynjs), 它们的主要特点是主成分2较大, 光合参数中Tr值最小; 第2类为广东乐昌(lcs)、湖北建始(js)、福建漳州(zz)、重庆彭水(ps)和广西兴安(xa), 它们的主要特点是主成分1、主成分2较相当, Pn、Tr、WUE值较大, 即光合速率和水分利用效率都较高; 第3类为剩余的6个种源, 它们的主要特点是主成分2较小、主成分1较大, 即光合较强, 水分利用效率较低。

|
图 1 14个香椿种源的二维主坐标分析图 Figure 1 Two-dimensional principal coordinate analysis diagram of 14 Toona sinensis provenances wd:甘肃武都种源; ynjs:云南建水种源; cd:湖南常德种源; xa:广西兴安种源; lcs:广东乐昌种源; js:湖北建始种源; ps:重庆彭水种源; zz:福建漳州种源; nn:广西南宁种源; ds:贵州独山种源; fh:湖南凤凰种源; ak:陕西安康种源; pa:四川蓬安种源; np:广西那坡种源。 |
香椿各个光合指标在种源间存在极显著差异, 与杨玉珍等[16]的研究相符。5个光合指标的变异系数变动幅度为4.42%~27.52%。其中, Gs变异系数最大, 变异幅度0.175~0.467 μmol·m-2·s-1, 说明香椿光合生理指标在种源间的差异是明显的。而Ci的变异系数最小, 为4.24%, 变异幅度247.856~292.717 μmol·mol-1, 说明该性状在种源间的差异较小。相关性分析结果表明, Pn与Gs、Tr、苗高极显著正相关, Pn与地径显著正相关, Gs与Ci、Tr、苗高极显著正相关, Ci与Tr显著正相关, Tr与苗高极显著正相关, 与地径显著正相关, 苗高与地径、羽叶数极显著正相关, 地径与羽叶数极显著正相关。相关性分析结果与杨玉珍等[16]的研究结果相符, 发现香椿的Pn值越大, 其WUE越小, 表现为光合越强的种源水分利用效率越低。总的趋势是偏南方的香椿种源Gs明显较偏北方的种源大, 这可能与南方水资源更丰富有关。广西那坡香椿种源的Tr值明显大于其他4个种源, 其次分别为重庆彭水、福建漳州、湖南凤凰、贵州独山。广西那坡、福建漳州种源的WUE值较低, 贵州独山、湖南凤凰种源的WUE值较高, 重庆彭水种源的WUE值居中, 说明偏南方的香椿种源水分利用效率较低, 偏北方的种源水分利用效率较高。这一结果与其他树种不完全相同, 比如黄连木Pistacia chinensis、猴樟Cinnamomum bodinieri、钩栗Castanopsis tibetana等[17-19]。
广西兴安、广西那坡、四川蓬安香椿种源的Pn值较大, 分别达到了19.513、17.641、16.543 μmol·m-2·s-1, 云南建水、湖南常德、甘肃武都种源的Pn较小, 分别为9.006、10.415、11.837 μmol·m-2·s-1。WUE最大的是广西兴安、广东乐昌种源, 为3.416、3.038 μmol·mol-1, 最小的是广西那坡种源, 为2.445 μmol·mol-1。可把14个香椿种源分为3类:第1类为甘肃武都(wd)、湖南常德(cd)、云南建水(ynjs)种源, 它们的主要特点是主成分2较大, 光合参数中Tr值最小; 第2类为广东乐昌(lcs)、湖北建始(js)、福建漳州(zz)、重庆彭水(ps)、广西兴安(xa)种源, 它们的主要特点是主成分1、主成分2较相当, Pn、Tr、WUE较大, 即光合速率和水分利用效率都较高; 第3类为剩余的6个种源, 它们的主要特点是主成分2较小、主成分1较大, 即光合作用较强, 水分利用效率较低。这一分类与武冲等[20]对麻楝Chukrasia tabularis种源的分类相似, 在实际生产中选择种源应做到因地制宜, 南方种植可不考虑水分利用效率, 宜选择南方种源, 北方种植则需综合指标更好的、适应性更全面的种源。
| [1] |
孙鸿有, 王鹏飞, 方炳法, 等. 香椿地理变异与种源选择[J]. 浙江林学院学报, 1992, 9(3): 4-12. (  0) 0) |
| [2] |
CHIA Y C, RAJBANSHI R, CALHOUN C, et al. Anti-neoplastic effects of gallic acid, a major component of Toona sinensis leaf extract, on oral squamous carcinoma cells[J]. Molecules, 2010, 15(11): 8377-8389. DOI:10.3390/molecules15118377 ( 0) |
| [3] |
ZHEN H, ZHANG Y, FANG Z, et al. Toona sinensis and Moschus decoction induced cell cycle arrest in human cervical carcinoma HeLa cells[J/OL]. Evid Based Complement Alternat Med, 2014, 2014: 1-8[2015-12-09]. http://dx.doi.org/10.1155/2014/121276.
( 0) |
| [4] |
WANG C C, TSAI Y J, HSIEH Y C, et al. The aqueous extract from Toona sinensis leaves inhibits microglia-mediated neuroinflammation[J]. Kaohsiung J Med Sci, 2014, 30(2): 73-81. DOI:10.1016/j.kjms.2013.09.012 ( 0) |
| [5] |
张友元, 夏玉芳, 许建初, 等. 香椿木材解剖构造及其物理力学性质[J]. 植物分类与资源学报, 2013, 35(5): 641-646. ( 0) |
| [6] |
骆嘉言, 林金国, 李大岔, 等. 香椿人工林和天然林木材材性的比较研究[J]. 西北林学院学报, 2003, 18(2): 77-79. DOI:10.3969/j.issn.1001-7461.2003.02.023 ( 0) |
| [7] |
范振富, 高瑞龙, 王杰铃. 香椿人工林和天然林木材纤维形态和化学成分比较研究[J]. 亚热带植物科学, 2003, 32(3): 35-37. DOI:10.3969/j.issn.1009-7791.2003.03.009 ( 0) |
| [8] |
范振富. 香椿速生丰产用材林栽培试验[J]. 林业科技开发, 2004, 18(5): 61-63. DOI:10.3969/j.issn.1000-8101.2004.05.023 ( 0) |
| [9] |
陈美珍. 香椿山地丰产林栽培技术研究[J]. 防护林科技, 2011, 103(4): 54-56. DOI:10.3969/j.issn.1005-5215.2011.04.019 ( 0) |
| [10] |
周小玲, 袁穗波, 李二平, 等. 香椿丰产栽培技术研究[J]. 中南林学院学报, 1997, 17(4): 64-69. ( 0) |
| [11] |
张玉洁. 香椿幼树光合作用及其影响因子研究[J]. 林业科学研究, 2002, 15(4): 432-436. DOI:10.3321/j.issn:1001-1498.2002.04.012 ( 0) |
| [12] |
杨玉珍, 张云霞, 彭方仁. 干旱胁迫对不同种源香椿苗木光合特性的影响[J]. 北京林业大学学报, 2011, 33(1): 44-48. DOI:10.3969/j.issn.1671-6116.2011.01.008 ( 0) |
| [13] |
郝明灼, 李群, 彭方仁, 等. 不同种源香椿芽菜品质分析及光合特性比较[J]. 林业科技开发, 2012, 26(4): 48-52. DOI:10.3969/j.issn.1000-8101.2012.04.012 ( 0) |
| [14] |
孟丽, 李德生, 茹丽叶, 等. Cd2+、Hg2+污染对香椿叶绿素含量和光合特性的影响[J]. 水土保持学报, 2013, 27(5): 240-243. DOI:10.3969/j.issn.1009-2242.2013.05.047 ( 0) |
| [15] |
孟生旺. 用主成份分析法进行多指标综合评价应注意的问题[J]. 统计研究, 1992, 48(4): 67-68. ( 0) |
| [16] |
杨玉珍, 贾遂民, 彭方仁. 不同种源香椿苗木光合特性比较[J]. 林业科技开发, 2008, 22(4): 62-64. DOI:10.3969/j.issn.1000-8101.2008.04.016 ( 0) |
| [17] |
董倩, 唐秀光, 王洁, 等. 不同种源黄连木光合参数比较及聚类分析[J]. 河北农业大学学报, 2012, 35(3): 58-62. ( 0) |
| [18] |
王娈, 韦小丽, 张怡. 不同地理种源猴樟苗期的光合特性与蒸腾特性[J]. 贵州农业科学, 2011, 39(6): 39-42. DOI:10.3969/j.issn.1001-3601.2011.06.011 ( 0) |
| [19] |
王佩兰, 许德禄, 张斌, 等. 钩栗种源间幼苗生长状况和光合特性比较研究[J]. 北方园艺, 2014, 38(5): 20-25. ( 0) |
| [20] |
武冲, 仲崇禄, 张勇, 等. 麻楝生长和光合作用参数种源变异分析[J]. 热带作物学报, 2014, 35(3): 509-514. DOI:10.3969/j.issn.1000-2561.2014.03.017 ( 0) |