 2016, Vol. 37
2016, Vol. 37



2. 华南农业大学 林学与风景园林学院/广东省森林植物种质创新与利用重点实验室, 广东 广州 510642;
3. 嘉应学院 生命科学学院, 广东 梅州 514015
2. College of Forestry and Landscape Architecture, Sourth China Agricultural University/Guangdong Key Laboratory for Innovative Development and Utilization of Forest Plant Germplasm, Guangzhou 510642, China;
3. School of Life Science, Jiaying University, Meizhou 514015, China
光合作用是绿色植物吸收和利用光能、水分和CO2, 制造有机物并释放氧气的过程[1], 是植物有机质合成、能量贮存与转化的基础[2], 已有大量研究证实光照是植物光合作用的主要限制因子[1-4]。光响应曲线是研究植物光合特性的重要手段之一, 通过光响应曲线数学模型可以计算出植物的最大光合速率、光补偿点、光饱和点、表观量子效率及暗呼吸速率等光响应特征参数, 这些参数可以一定程度反映植物的光合作用潜力, 也可以阐明植物光合产物积累与外界光照条件的联系[3-4]。
刨花润楠Machilus pauhoi, 又名刨花楠、刨花(广东)、黏柴(福州), 属樟科(Lauraceae)润楠属Machilus常绿大乔木。刨花润楠是一个亚热带树种, 喜阴耐湿, 产于长江以南东南各省海拔300~1 200 m, 生于山坡及沟谷, 与甜槠Castanopsis eyrei、木荷Schima superba、桃Amygdlu spersica等混生成小片纯林。刨花润楠生长迅速, 树干通直, 出材量大, 从叶中提取精油, 不仅是优良的天然香料, 且精油中所含薁化物(Azulenes), 具有独特的药用价值, 其茎是制作药物“白楠木”的原料; 树皮含树脂、橡胶; 种子含油率高, 是优良的工业润滑油, 可供制皂及制蜡用; 刨花润楠树体高大, 树形美观, 枝繁叶茂, 四季常青, 嫩叶呈粉红色或红棕色, 是优良的庭院观赏、园林绿化树种。由于刨花润楠全身是宝, 因此是一种极具经济价值和开发前景的阔叶树种[5-7]。目前对刨花润楠的研究主要集中于育苗和繁殖技术等方面, 生理生态方面的研究还很少, 尤其缺乏对刨花润楠光合特性的认识。就目前的研究来看, 关于刨花润楠不同种源之间光合特性的对比研究鲜见报道。但是, 在刨花润楠的造林过程中, 立地因子筛选、造林密度控制等方面均与刨花润楠的光合利用和适应能力密切相关, 因此, 科学认识刨花润楠的光合能力以及对光照强度的响应规律很有必要。本文通过对不同种源的刨花润楠光响应曲线特征进行对比, 分析其光合特性的种源变异, 阐明刨花润楠种源之间光合生产能力和光适应能力, 并为刨花润楠良种选育与造林提供依据。
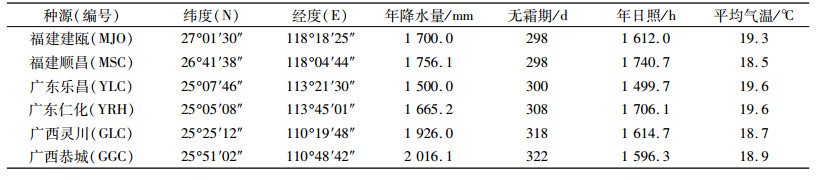
1 材料与方法 1.1 试验材料和试验地选取位于树冠中部外层、叶龄相近的功能叶片进行测量。试验选取6个不同种源的刨花润楠种子培育的1年生苗木, 其分别为福建建瓯种源(MJO)、福建顺昌种源(MSC)、广东乐昌种源(YLC)、广东仁化种源(YRH)、广西灵川种源(GLC)、广西恭城种源(GGC), 各种源采种地的地理位置与气候因子见表 1。试验地点位于广州市华南农业大学林学院教学科研基地, 地理位置E 113°37′, N 23°16′, 海拔42.3 m, 属于南亚热带季风气候区, 平均气温21.9 ℃, 最冷月平均气温11.0 ℃, 最热月平均气温28.7 ℃, 最低气温为-2.6 ℃, 最高气温为39.3 ℃。平均年降水量1 696.5 mm, 年降水150 d左右, 降雨多集中于4—6月份。年平均相对湿度68%, 平均霜期每年1 d, 年平均日照1 800 h。
|
|
表 1 刨花润楠种源地理位置与气候因子 Table 1 Geographic locations and climatic information of the seed collection sites of different Machilus pauhoi provenances |
2014年8月下旬, 选择晴朗天气, 使用Li-6400XT便携式光合作用测定仪测定叶片各光合生理指标。测定时间为09:00—11:30。叶温设置为30 ℃, 测量时采用开放气路, 大气CO2设置为400 μmol·mol-1, CO2匹配值为20 μmol·mol-1。使用LI-6400-02B红蓝光源, 测定系统调节光合有效辐射在0~2 000 μmol·m-2·s-1范围内, 叶室内光量子通量密度(PPFD)的梯度设置为2 000、1 800、1 500、1 200、1 000、800、600、400、200、150、100、50、25、0 μmol·m-2·s-1。选取位于树冠中部外层、叶龄相近的功能叶片(从顶部开始的第3~5片)进行测量。每种源测定3株, 每株测3片叶, 重复5次, 取其平均值进行分析。测定光合生理指标包括:净光合速率(Pn)、气孔导度(Gs)、蒸腾速率(Tr)、胞间CO2浓度(Ci)。
1.3 光响应曲线分析方法利用Excel以及SPSS19.0对试验数据进行整理及统计分析。用Origin Pro v8.0软件作图。
根据基于光合作用的羧化和电子传递2个基本过程描述光合作用与生物化学之间关系的叶片Farquhar模型[8], 采用迭代法对光响应曲线进行拟合, 并对所得数据进行非线性回归分析, 计算最大净光合速率(Pnmax)、光响应曲线曲角(K)和暗呼吸速率(Rd)[9-11]。同时, 将200 μmol·m-2·s-1以下光量子通量密度(PPFD)的净光合速率进行直线回归分析, 求出表观量子效率(Apparent quantum yield, φ)、光饱合点(Light saturation point), 光补偿点(Light compensation point)[9]。
Farquhar模型如下:
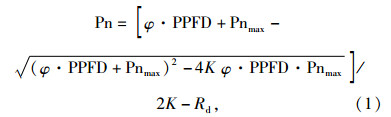
|
(1) |
其中:Rd为暗呼吸速率; Pn、PPFD、Pnmax和Rd的单位均为μmol·m-2·s-1。
在拟合中分别设定初始值如下:K=0.5, Pnmax=30 μmol·m-2·s-1, φ=0.05, Rd=2 μmol·m-2·s-1[12]; 限制值设定为K≤1, Pnmax≤50 μmol·m-2·s-1, φ≤0.125, Rd不设定限制值。
将小于200 μmol-1.m-2.s-1的PPFD与Pn进行直线回归, 可按下式计算出光补偿点、光饱合点和φ等参数[13]:
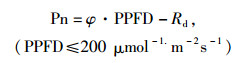
|
(2) |
当Pn为0时, PPFD为光补偿点; 当Pn为Pnmax时, PPFD为光饱合点。
2 结果与分析 2.1 不同种源刨花润楠光合生理指标对光强的响应如图 1A所示, 6个种源刨花润楠净光合速率随着光量子通量密度的增强而增大, 上升趋势基本相同。在PPFD为0~200 μmol·m-2·s-1范围内, 净光合速率随着PPFD的增强呈现迅速上升的变化趋势, 此后, 随着PPFD的增强, 各种源净光合速率的增加略为缓慢, 当PPFD到达一定强度后, 各种源刨花润楠净光合速率逐渐趋于与PPFD轴平行。不同种源刨花润楠叶片的气孔导度均随PPFD的增强而增大(图 1B), 而胞间CO2浓度则总体呈现出随PPFD增大先迅速减小后保持不变的变化趋势(图 1C)。就蒸腾速率而言, 由图 1D可知, 6个种源刨花润楠蒸腾速率对PPFD的响应曲线与气孔导度的响应曲线较为相似, 也表现出随着PPFD增强而增大的变化趋势。总体而言, 初步分析结果显示6个种源刨花润楠各光合生理指标随PPFD的变化表现出了相似的变化规律。

|
图 1 刨花润楠6个种源叶片净光合速率(Pn)、气孔导度(Gs)、胞间CO2浓度(Ci)和蒸腾速率(Tr)对光量子通量密度(PPFD)的响应 Figure 1 Responses of photosynthetic rate (Pn), stomatal conductance (Gs), intercellular CO2 concentration (Ci) and transpiration rate (Tr) to photosynthetic photon flux density (PPFD) in leaves of six Machilus pauhoi provenances |
在植物光合作用中, 最大净光合速率愈大, 表明植物利用光能的潜在能力愈强[14]。由表 2可以看出, 6个刨花润楠不同种源间最大净光合速率的变化幅度为5.32~12.20 μmol·m-2·s-1, 极差为6.88 μmol·m-2·s-1, 变异系数为29.77 %。各种源最大净光合速率从大到小依次为广西恭城、广东仁化、广西灵川、广东乐昌、福建顺昌、福建建瓯。福建2个种源的最大净光合速率显著低于另外4个种源。从总体上看, 光合作用潜在能力大小依次为广西种源、广东种源、福建种源。
|
|
表 2 刨花润楠不同种源的光响应参数1) Table 2 The light response parameters of different Machilus pauhoi provenances |
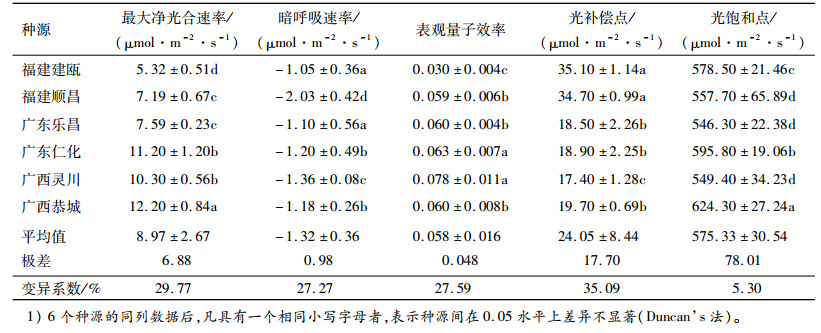
植物暗呼吸是一个消耗参数, 主要受植物生长状态和温度的影响。暗呼吸速率越小, 表明其消耗能量越多。从表 2可以看出, 不同种源刨花润楠的暗呼吸速率差异明显, 变幅在-2.03 ~-1.05 μmol·m-2·s-1, 极差为0.98 μmol·m-2·s-1, 变异系数为27.27 %。福建顺昌种源的暗呼吸速率最小, 表明暗呼吸作用较强, 消耗能量较多, 而福建建瓯暗呼吸速率最大, 表明福建建瓯暗呼吸作用较弱, 消耗能量较少。
2.2.2 表观量子效率6个刨花润楠种源的表观量子效率变幅0.030~0.078, 极差为0.048, 变异系数为27.59 %(表 2)。广西灵川种源的表观量子效率为0.078, 显著高于其他种源, 说明广西灵川种源刨花润楠利用弱光的能力较强; 而福建建瓯种源的刨花润楠则相反, 表观量子效率远小于其他5个种源, 不及广西灵川种源的一半, 说明福建建瓯种源利用弱光能力较弱。表观量子效率高, 说明能够有效地利用弱光, 因此从生产的角度来说, 更适合于密植或营造混交林。由于广西灵川种源刨花润楠的表观量子效率较高, 因此, 使用广西灵川种源造林, 可适度密植或营造混交林, 以提高林地的木材产量。
2.2.3 光补偿点和光饱和点由表 2可知, 6个刨花润楠种源的光补偿点的差异较大, 变化范围为17.40~35.10 μmol·m-2·s-1, 各种源光补偿点从大到小依次为福建建瓯、福建顺昌、广西恭城、广东仁化、广东乐昌、广西灵川, 极差为17.70μmol·m-2·s-1, 变异系数为35.09 %。福建2个种源的光补偿点显著高于其他2个省份种源, 说明福建种源利用弱光能力较其他4个种源弱, 广西灵川种源利用弱光能力最强, 与2.2.2中表观量子效率得出的福建2个种源利用弱光能力弱, 而广西灵川利用弱光能力强的结果一致。
刨花润楠不同种源光饱和点变化幅度为546.30~624.30 μmol·m-2·s-1, 最大的为广西恭城种源, 其次为广东仁化、福建建瓯、福建顺昌和广西灵川种源, 最小为广东乐昌种源, 极差为78.01μmol·m-2·s-1, 变异系数为5.30 %。不同种源刨花润楠光饱和点的结果表明, 广西恭城种源对强光的利用效率相对较高, 而广东乐昌种源对强光的利用效率相对较低。在6个不同种源中, 光饱和点与光补偿点之间的差值最大的为广西恭城种源, 为604.60 μmol·m-2·s-1, 说明广西恭城种源对光的适应性较其他5个种源强, 广东仁化种源次之, 而福建顺昌种源的光饱和点与光补偿点间差值最小, 说明福建顺昌种源的刨花润楠对光适应性最弱。
3 讨论与结论净光合速率的光响应曲线反映光合速率随光强改变的变化规律[15]。本研究结果表明, 参试的6个不同种源刨花润楠光合作用光响应曲线呈现出相似的变化趋势, 即净光合速率随着光量子通量密度(PPFD)的增强而增大, 符合光合速率随光强改变的变化规律; 但6个不同种源刨花润楠最大净光合速率的变化幅度为5.32~12.20 μmol·m-2·s-1, 整体表现光合速率偏低, 不同种源光合速率高低排序依次为广西恭城、广东仁化、广西灵川、广东乐昌、福建顺昌、福建建瓯, 初步反映广西恭城种源刨花润楠的生物质生产力较突出。
气孔是植物对大气进行水汽和CO2交换的通道, 可以反映树木自身的水分消耗能力和适应外界环境的能力, 其开闭程度对蒸腾作用、光合作用具有重要调控作用, 关系到植物的水分消耗和木材的形成[16]。光量子通量密度的增强导致光合速率增大, 消耗CO2的量也增大; 叶片通过调节气孔来弥补CO2的消耗, 增大气孔导度来增加外界CO2向叶肉细胞内的扩散[17]。本研究结果显示, 参试的6个不同种源刨花润楠叶片的气孔导度均随PPFD的增强而增大, 广东仁化种源、广西灵川种源和广西恭城种源气孔导度上升的趋势比较平缓, 但气孔导度较大, 且CO2浓度较大, 净光合速率也较高。可初步得出上述3个种源的光合作用相对较强。
水分是光合作用合成有机物所需的主要原料之一, 蒸腾作用为植物所需的外部矿质元素和水分的运输产生动力, 因此, 蒸腾速率与光合速率关系密切。通常情况下蒸腾速率随着PPFD的增强而增强。本研究中6个种源刨花润楠蒸腾速率对光强的响应曲线与气孔导度的响应曲线较为相似, 都表现出随着光强增强而增大的变化趋势。
光补偿点和光饱和点是衡量植物利用光强能力的重要指标[18]。一般地讲, 光补偿点越低的植物利用弱光能力越强, 而光饱和点高的植物, 能更有效地利用全日照的强光, 光饱和点高的植物生长较快[19-21]。本研究中, 广西灵川种源光补偿点最低, 说明广西灵川种源刨花润楠利用弱光能力最强, 同时广西恭城种源光饱和点最大, 对强光的利用效率相对较高。在6个不同种源刨花润楠中, 光饱和点与光补偿点之间的差值最大的为广西恭城种源, 说明广西恭城种源对光的适应性较其他5个种源强; 但从研究结果也可看出, 6个不同种源刨花润楠的光补偿点和光饱和点都不高, 综合分析各种源的光响应曲线可知, 参试的6个种源刨花润楠幼苗均具有一定的耐阴性。耐阴性是植物的一种重要性状, 对耐阴性的分析和比较对以后该树种在林分的营造中起到一定的作用。
表观量子效率是植物对CO2同化的表观量子效率, 衡量植物光合作用对光能的利用效率, 其数值越高, 叶片转化光能的效率也相应越高, 植物吸收与转换光能的色素蛋白复合体可能较多, 同时利用弱光的能力强[22-24]。本研究中, 广西灵川种源刨花润楠的表观量子效率较高, 因此, 从理论推断, 在实际造林中, 可适度密植或营造混交林, 以提高林地木材产量[25]。
综上所述, 参试的6个刨花润楠种源中, 广西恭城种源刨花润楠利用弱光、强光能力和对光适应能力最强, 光合潜力较大, 具有较好的生长潜力。
| [1] |
蒋冬月, 钱永强, 费英杰, 等. 柳属植物光合-光响应曲线模型拟合[J]. 核农学报, 2015, 29(1): 169-177. (  0) 0) |
| [2] |
黄雅丽, 梁士楚, 韦宇静, 等. 巨尾桉与黧蒴栲、红锥光响应曲线的比较研究[J]. 广西科学院学报, 2014, 30(4): 222-228. DOI:10.3969/j.issn.1002-7378.2014.04.002 ( 0) |
| [3] |
邱权, 潘昕, 何茜, 等. 华南地区3种苗木生长旺盛时期光合特性及蒸腾耗水日变化规律的比较[J]. 华南农业大学学报, 2012, 33(4): 524-528, 549. ( 0) |
| [4] |
俞新妥, 卢建煌. 不同种源马尾松光合能力的比较研究[J]. 福建林学院学报, 1991, 11(2): 131-135. ( 0) |
| [5] |
吴振伙, 吴兆平, 全尚龙. 刨花润楠综合利用价值及其育苗技术[J]. 现代农业科技, 2008(20): 55. DOI:10.3969/j.issn.1007-5739.2008.20.035 ( 0) |
| [6] |
胡希华. 刨花润楠的优良特性及育苗栽培技术[J]. 湖南林业科技, 2006, 33(1): 65-66. DOI:10.3969/j.issn.1003-5710.2006.01.025 ( 0) |
| [7] |
钟智群, 谭梓峰, 杨志玲, 等. 刨花楠生长发育特点及开发前景分析[J]. 湖南林业科技, 1997, 24(2): 50-51. ( 0) |
| [8] |
FARQUHAR G D, VON CAEMMERER S, BERRY J A. Models of photosynthesis[J]. Plant Physiol, 2001, 125(1): 42-45. DOI:10.1104/pp.125.1.42 ( 0) |
| [9] |
WALKER D A, JARVIS P G, FARQUHAR G D, et al. Automated measurement of leaf photosynthetic O2 evolution as a function of photon flux density[J]. Philos Trans R Soc London:Ser B:Biol Sci, 1989, 323(1216): 313-326. DOI:10.1098/rstb.1989.0013 ( 0) |
| [10] |
蒋高明, 林光辉, MARINOB D V. 美国生物圈二号内生长在高CO2浓度下的10种植物气孔导度、蒸腾速率及水分利用效率的变化[J]. 植物学报, 1997, 39(6): 546-553. ( 0) |
| [11] |
温达志, 周国逸, 张德强, 等. 四种禾本科牧草植物蒸腾速率与水分利用效率的比较[J]. 热带亚热带植物学报, 2000, 8(增刊Ⅲ): 67-76. ( 0) |
| [12] |
HERRICK J D, THOMAS R B. Effects of CO2 enrichment on the photosynthetic light response of sun and shade leaves of canopy sweetgum trees (Liquidambar styraciflua) in a forest ecosystem[J]. Tree Physiol, 1999, 19(12): 779-786. DOI:10.1093/treephys/19.12.779 ( 0) |
| [13] |
RICHARDSON A D, BERLYN G P. Spectral reflectance and photosynthetic properties of Betula papyrifera (Betulaceae) leaves along an elevational gradient on Mt.Mansfield, Vermont, USA[J]. Am J Bot, 2002, 89(1): 88-94. DOI:10.3732/ajb.89.1.88 ( 0) |
| [14] |
许大全. 光合作用效率[J]. 植物生理学通讯, 1988(5): 1-7. ( 0) |
| [15] |
滕建国, 高长启, 林玉梅, 等. 加拿大黄桦幼苗光合特性的研究[J]. 吉林林业科技, 2006, 35(2): 5-7. DOI:10.3969/j.issn.1005-7129.2006.02.002 ( 0) |
| [16] |
何茜, 李吉跃, 姜枫. 毛白杨抗旱节水优良无性系评价与筛选[D]. 北京: 北京林业大学, 2008.
( 0) |
| [17] |
张力文, 钟国成, 张利, 等. 3种鼠尾草属植物光合作用-光响应特性研究[J]. 草业学报, 2012, 21(2): 70-76. ( 0) |
| [18] |
金则新, 柯世省. 云锦杜鹃叶片光合作用日变化特征[J]. 植物研究, 2004, 24(4): 447-452. DOI:10.3969/j.issn.1673-5102.2004.04.021 ( 0) |
| [19] |
申文辉, 李志辉, 彭玉华, 等. 格木不同种源光合作用光响应分析研究[J]. 中南林业科技大学学报, 2014, 34(6): 13-18. DOI:10.3969/j.issn.1673-923X.2014.06.004 ( 0) |
| [20] |
宋庆安, 李昌珠, 童方平, 等. 光皮树优良无性系光合生理特性对光强的响应[J]. 湖南林业科技, 2008, 35(6): 3-6. DOI:10.3969/j.issn.1003-5710.2008.06.002 ( 0) |
| [21] |
郑元. 刺槐光合生理特征与固碳能力研究[D]. 咸阳: 西北农林科技大学, 2010. http://cpfd.cnki.com.cn/Article/CPFDTOTAL-ZGLX200911004032.htm
( 0) |
| [22] |
胡婧楠. 三种楠木光合生理特性及叶子养分年变化的比较研究[D]. 合肥: 安徽农业大学, 2010.
( 0) |
| [23] |
许大全. 叶片表观光合量子效率的测定[M]//上海市植物生理学会, 中科院植物生理研究所. 现代植物生理学实验指南. 北京: 科学出版社, 1999.
( 0) |
| [24] |
许大全. 光合作用效率[M]. 上海: 上海科学技术出版社, 2002.
( 0) |
| [25] |
郭江, 郭新宇, 王纪华, 等. 不同株型玉米光响应曲线的特征参数研究[J]. 西北植物学报, 2005, 25(8): 1612-1617. DOI:10.3321/j.issn:1000-4025.2005.08.021 ( 0) |