 2016, Vol. 37
2016, Vol. 37



任豆Zenia insignis又名翅荚木、任木,为苏木科任豆属单种属植物[1],是落叶大乔木,列为国家二级重点保护植物[2]。任豆是印度-马来西亚-越南(或中南半岛)至华南(或西南)区域分布的植物,在我国主要分布于中亚热带与南亚热带之间的广西、广东、云南、湖南、贵州等地[3],是华南石灰岩地区的特有种。任豆用途极为广泛,它的抗旱、抗热能力强,已经成为石质山地造林、植被恢复重建的首选乡土树种之一[4]。不仅可以涵养水源、保持水土,调节气候,改善种植地的生态环境,重建退化的森林生态系统[5],还具有抗二氧化硫的作用[6]。木材纤维特性好,是理想的人造板及纸张生产原料[7-8]。叶片营养丰富,适口性强,刈割性好,是值得开发利用的木本饲料树种[9]。此外,任豆还是良好的薪炭树种和良好的蜜源树种。任豆作为我国珍稀树种,也是值得推广利用的优良树种,对其遗传多样性的评价和遗传改良是很有必要的。本文旨在通过分析任豆不同种源种子性状地理变异,揭示任豆种子表型性状地理变异及其模式。
1 材料与方法 1.1 供试种源在2013—2014年间,从任豆主要分布区的5省21县(市、区)采集种子,每个采种点的采种母树为20~30株,要求母树树型通直圆满、生长健壮、没有受病虫害侵袭,达到正常开花结实年龄。母树间直线距离要求大于100 m。同一采种点母树种子均匀混合作为供试种源材料。各种源编号、采种点地理位置、主要气候因子状况见表 1。
|
|
表 1 任豆种源采种点地理位置和气候因子状况 Table 1 Geographic location and climatic information of the seed collection sites of different Zenia insignis provenances |
每个种源随机抽取纯净种子30粒,使用游标卡尺测量每粒种子的长度、宽度、厚度(种子最厚处),精确到0.01 mm。每个种源随机抽取纯净种子100粒,用电子天平称量种子质量,重复抽取5次,取平均值,精确到0.01 g。
1.3 数据统计分析采用Excel2007软件整理数据。方差分析、相关分析、偏相关与聚类分析使用Spss19.0软件。聚类分析和偏相关分析的数据为种源平均值,并对数据采用“全局-1~1”标准化处理。最后,采用欧氏距离离差平方和法进行聚类分析。
为了解各性状在种源间的差异和遗传稳定性,计算变异系数(CV)和重复力(R):
CV=SD/MIN×100%,
R=1-1/F,
式中,SD为性状标准差,MIN为性状平均数, F为方差分析中种源间与种源内的均方比。
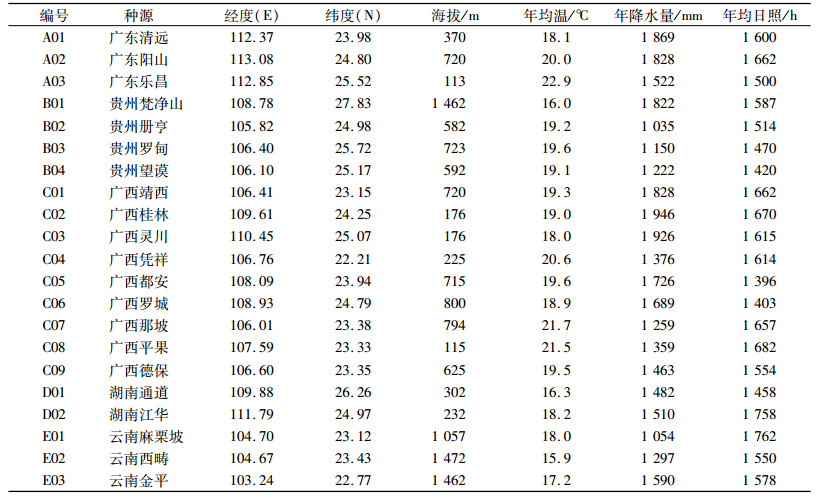
2 结果与分析 2.1 不同种源种子性状变异分析任豆种源间种子性状的差异分析结果(表 2)显示,任豆种子各性状在种源间存在一定程度的差异,各性状的变异幅度也存在差异,6个种子性状间变异系数变动幅度在5.76%~10.16%。其中,种子厚度的变异系数最大,达到10.16%,变异幅度1.24~2.84 mm,说明该性状在种源间的差异更明显。而种子宽度变异系数最小,为5.76%,变异幅度3.45~6.49 mm,说明该性状在种源间的差异较小。
|
|
表 2 任豆种源间种子性状变异统计 Table 2 Summary of variations in seed traits of different Zenia insignis provenances |
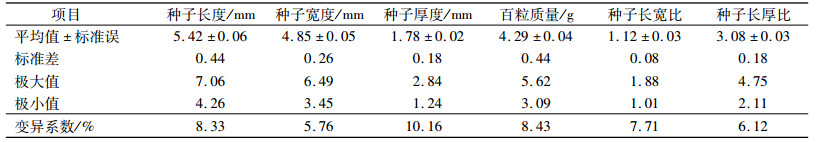
由表型性状方差分析结果(表 3)可以看出,任豆种源间在种子长度、宽度、厚度、长宽比、长厚比及百粒质量上都存在极显著差异。F值由大到小依次为种子长度(23.988)、厚度(14.409)、宽度(13.304)、长厚比(11.469)、百粒质量(6.907)和长宽比(6.333)。6个性状的重复力变动范围为84.2%~95.8%。其中,种子长度的重复力最高,达到95.8%,其次是种子厚度、宽度和长厚比,这3个性状的重复力均高达90%以上。种子长宽比的重复力最低,但也达到84.2%。表明任豆种源间在这6个性状上的差异均具有一定的稳定性,可以作为任豆种子性状遗传多样性评价和地理变异研究的主要性状。
|
|
表 3 任豆不同种源种子表型性状方差分析 Table 3 Analysis of variance of seed traits of different Zenia insignis provenances |
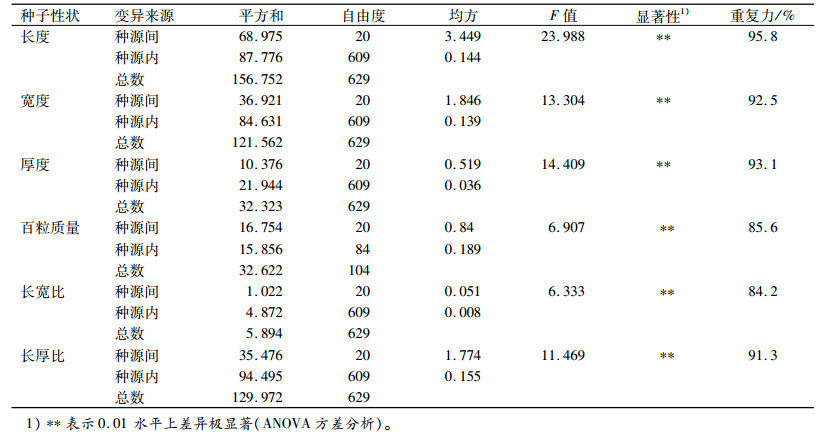
种子性状和采种点地理因子偏相关分析结果(表 4)表明,种子6个性状均与纬度呈负相关,其中,种子厚度和百粒质量与纬度呈显著负相关,这说明采种点纬度从低到高,任豆种子逐渐变薄,变轻。种子性状与经度均呈负相关,但没有达到显著水平。种子性状都与海拔呈负相关,其中,种子长宽比与海拔相关性达到显著水平,表明种子形状随着海拔的升高逐渐变得扁平,由圆形逐渐变化为椭圆形。整体来看,任豆种子形态性状存在显著的产地差异,具有一定的纬度向变异趋势,西南山区的任豆种子(云南、贵州、广西种源)比东部(湖南、广东种源)种粒大。
|
|
表 4 任豆种子表型性状与采种点地理因子的偏相关分析1) Table 4 Partial correlation coefficients between geographic factors of collection sites and Zenia insignis seedtraits |
为揭示任豆种子性状变异的气候生态学基础,对各种源的6个种子性状与采种点3个气候因子(年均温、年降水量、年均日照)作了偏相关分析,结果(表 5)表明,种子长度、宽度、长宽比、长厚比、百粒质量与年平均温度呈显著负相关关系,6种性状与降水量呈正相关关系,其中,种子百粒质量与年均降水量的相关关系达到显著水平。采种点日照时数与种子性状相关关系未达到显著水平。
|
|
表 5 任豆种子表型性状与采种点气候因子的偏相关分析1) Table 5 Partial correlation coefficients between climate factors of collection sites and Zenia insignis seed traits |

由此可以看出,在气候因子中,气温的影响最大,其次是降水量。年平均温度越高的采种点,任豆种子的长度和宽度越小,种子长宽比、长厚比也会越小,种子逐渐呈现圆形并且厚度降低,且百粒质量越轻。表明任豆种子的大小和重量主要受分布地区气温的影响,随年均温增大而变小。年平均降水量越大,种子的百粒质量也会随之变得越来越大。
2.4 任豆种子各性状相关性分析各种源种子性状之间的相关性分析结果(表 6)表明,种子长度与种子宽度、长宽比、长厚比、百粒质量都呈现极显著正相关,相关系数分别为0.921、0.852、0.758、0.889,说明了任豆种子的长度与其余各个性状相关关系更紧密,种子越长,种粒越大。种子宽度与种子长厚比、百粒质量呈极显著正相关,相关系数分别达到0.744和0.865;种子厚度与种子百粒质量呈现显著正相关,表明种子越厚,种子质量越大;种子长宽比和种子的百粒质量也呈现显著正相关关系,表示种子性状越接近椭圆形,种子质量越大。
|
|
表 6 任豆种子表型性状相关分析1) Table 6 The correlation analysis on Zenia insignis seed traits |

根据种子6个性状,采用欧式距离离差平方和法对21个种源进行聚类分析,结果见图 1。当欧式距离为7(图 1虚线所示)时,21个种源的种子可以划分为5个类群。第1类群为B03、C03、C04、C05、C06、C07、和C08种源,包括广西的6个种源和贵州的1个种源,处于任豆分布区域的中西部,这类种源种粒大。第2类为C01、C02、C09、B02和B04种源,包括了3个广西的种源和2个贵州的种源,从地理位置上看,主要处于广西和贵州的交界处,其种子大小和质量居中,种子长宽比、种子长厚比也居中。第3类为A01、A02、D01和D02种源,地理位置处于广东和湖南的交界处,其种子颗粒较小,较薄,质量较轻。第4类为A03种源,这个种源处于任豆分布区域最东部的广东乐昌,它的种子较小,质量也稍低于平均值。第5类为B01、E01、E02和E03种源,即3个云南种源和1个贵州种源,其种子性状指标高于平均值。从上可知,聚类结果总体上具有明显的地域性。

|
图 1 任豆种源种子性状聚类结果 Figure 1 Dendrogram of Zenia insignis provenances based on seed traits C04:广西凭祥种源;C08:广西平果种源;C07:广西那坡种源;C06:广西罗城种源;C03:广西灵川种源;C05:广西都安种源;B03:贵州罗甸种源;C01:广西靖西种源;C02:广西桂林种源;B02:贵州册亨种源;B04:贵州望谟种源;C09:广西德保种源;D01:湖南通道种源;D02:湖南江华种源;A01:广东清远种源;A02:广东阳山种源;A03:广东乐昌种源;E02:云南西畴种源;E03:云南金平种源;E01:云南麻栗坡种源;B01:贵州梵净山种源。 |
较高的形态稳定性使得树木种子可以作为分类的重要依据[10],对树种遗传多样性研究具有重要价值。任豆作为我国西南石灰岩地区特有树种,其自然分布由云贵高原一直向东延伸到广东、湖南。生存环境特殊,由喀斯特地貌造成的群落分布地带支离破碎,地理隔离使得不同的地理种源长期在各自不同区域中生长繁育,有可能促使某些性状在不同种源之间产生遗传分化。本研究共收集5个省21个种源,对6个性状观测与分析的结果表明,6个种子性状间变异系数变动幅度5.60%~11.16%,且在种源间都存在极显著差异,说明任豆种子性状在种源间存在丰富的变异与多样性。6个性状的重复力变动范围为84.2%~95.8%。其中,种子长度、宽度、厚度和百粒质量这4个性状的重复力高达90%以上。由此可以说明,任豆种子的6个性状差异均主要由遗传因子控制,可作为任豆性状遗传多样性评价和地理变异研究的主要依据。
任豆种子形态特征(长度、宽度、厚度、长宽比、长厚比及百粒质量)种源间存在显著差异,种源效应明显。种子大小具有一定的纬度向变异,百粒质量受经、纬度共同影响。产于任豆分布区西南部的种子(贵州以及广西)较东北部(湖南、广东)种源的种子颗粒大且饱满。种子的形态自西向东由长圆柱体慢慢向短粗圆柱体变化。长度、宽度、长宽比等均呈随纬度变化的趋势,任豆地理分布越靠南,种子形态越接近长圆柱体。任豆种子百粒质量与种子长度、宽度以及厚度相关,表明种子的大小直接影响种子的质量。任豆种子大小与温度呈极显著负相关,与降水量呈正相关,这表明产地的年均温是影响种子大小的主要气候因子,而产地降雨量是影响百粒质量差异的主要气候因子。
任豆种子变异与地理相关研究报道少,仅有何小勇[11]对任豆9个种源种子作了观测,研究表明,产于贵州的种子较大,湖南和广东北部的种子较小,其中贵州兴义的种子长度、宽度及种子千粒质量显著大于其他地区的种子,最小的是广东种源的种子。种子的大小与经纬度呈现负相关关系,与本研究的结果相近。在其他树种的地理变异研究中也曾发现有类似的现象,这种现象可能与树种对不同分布区域的局部地理、气候条件适应有关。如梁有旺[12]发现香椿Toona sinensis种子的大小与经纬度负相关,百粒质量与海拔正相关,并与年均温、经纬度负相关,本研究结果中百粒质量与纬度、年均降雨量的相关性与之相符。程诗明等[13]对苦楝Melia azedarach 729个家系的18个表型性状因子与5个地理气候因子的相关分析研究发现,具有明显的性状东北、西南方向趋势变异的特点:长度由南到北逐渐变大;宽度由东向西逐渐变大;南部、东部分布的家系比北部、西部的家系生长快,种粒小。王家源等[14]对10个种源苦楝种子生物学特性的研究结果表明,苦楝种子由北往南体积和质量有变小的趋势,种子形状也逐渐由长圆柱形向圆球形改变。这些研究所揭示的种子性状地理变异趋势与本研究得到结论是基本一致的。
根据任豆6个性状的聚类分析,可将21个种源分为5大类,不同类群种子性状差异明显,广西中西部低山地区以及云南和贵州的种源种子个体及质量均较大,而广东和湖南交界处的种粒较小,且较轻,表现由西南往东北向的种子变小变圆的趋势。任豆种源类群划分结果具有明显的地理区域性,能够为任豆种质资源的区划提供一定的依据。
| [1] |
CHEN W Y. A new genus in the Chinese flora[J]. Sunyatsenia, 1946, 6(34): 195-198. (  0) 0) |
| [2] |
傅立国. 中国植物红皮书:稀有濒危植物:第1册[M]. 北京: 科学出版社, 1992.
( 0) |
| [3] |
陈永密. 珍稀树种:翅荚木[M]. 福州: 福建教育出版社, 1989.
( 0) |
| [4] |
侯伦灯, 李玉曹, 李平宇, 等. 任豆树综合利用研究[J]. 林业科学, 2010, 37(3): 139-143. ( 0) |
| [5] |
唐兰芳. 石灰岩地区造林绿化好树种:任豆[J]. 广东林业科技, 1998, 14(3): 34-37. ( 0) |
| [6] |
KIM S E, KIM H S, HONGY S, et al. Sesquiterpene esters from Celastrus orbiculatus and their structure-activity relationship on the modulation of multidrug resistance[J]. J Nat Prod, 1999, 62(5): 697-700. DOI:10.1021/np9804379 ( 0) |
| [7] |
周志春, 李建民, 陈炳星, 等. 几种亚热带速生乡土阔叶树种的制浆特性评价[J]. 中国造纸, 2003, 22(2): 7-11. ( 0) |
| [8] |
侯伦灯, 李玉蕾, 李平宇, 等. 任豆树综合利用研究[J]. 林业科学, 2001, 37(3): 139-143. DOI:10.3321/j.issn:1001-7488.2001.03.022 ( 0) |
| [9] |
范霭萱, 梁兆彦, 宋喜宣. 可开发的木本饲料[J]. 广西畜牧兽医, 1995, 11(2): 20-23. ( 0) |
| [10] |
张鹏, 沈海龙. 花曲柳优树种子性状变异及其子代苗期生长表现[J]. 林业科技开发, 2013, 27(2): 18-19. DOI:10.3969/j.issn.1000-8101.2013.02.005 ( 0) |
| [11] |
何小勇. 翅荚木种源遗传多样性研究[D]. 长沙: 中南林业科技大学, 2007.
( 0) |
| [12] |
梁有旺. 不同种源香椿种子及苗木差异性分析[D]. 南京: 南京林业大学, 2006.
( 0) |
| [13] |
程诗明, 顾万春. 苦楝遗传资源学研究进展及其展望[J]. 浙江林业科技, 2007, 27(2): 64-69. DOI:10.3969/j.issn.1001-3776.2007.02.017 ( 0) |
| [14] |
王家源, 郭杰, 喻方圆. 不同种源苦楝种子生物学特性差异[J]. 南京林业大学学报(自然科学版), 2013, 37(1): 49-54. ( 0) |