 2016, Vol. 37
2016, Vol. 37



苦楝Melia azedarach为楝科Meliaceae楝属Melia落叶乔木,是速生、优质的多用途树种。其木材轻软坚韧、纹理粗而美、有光泽、易加工、抗白蚁、防虫蛀和耐腐,可供家具、建筑、农具、船舶和乐器使用[1]。苦楝的根、皮、花、果均可入药,其根、皮、果实中提取出苦楝素,可制作成植物农药。苦楝也是城市绿化树种,具较强的耐烟尘、抗二氧化硫等优良性状,适宜矿区绿化[2-3]。苦楝分布广泛,自南向北横跨热带、亚热带、温带。从我国北方的河北保定、山西运城和甘肃陇南到南方的海南三亚,从东部的台湾、福建到西部的四川和云南,中国近1/3的陆地面积都有苦楝生长[4]。由于生态环境条件的差异大,长期的地理隔离和自然选择,可能会形成不同的地理生态类型。
国内有学者对苦楝开展过一些种源试验和地理变异研究。教忠意等[5]对江苏、山东、河南、福建等4省采集的20个苦楝家系1年生实生苗进行性状测定和分析;陈羡德等[6]对福建省漳平市的4年生苦楝种源试验林进行调查分析;陈丽君等[7]对17个省份的70个苦楝种源的果核和种子的18个性状进行了观测和分析。但对苦楝生长性状地理变异研究所涉及的种源较少,且并未覆盖全分布区。
本研究在19个省(自治区)搜集苦楝种源53个,采种区覆盖了苦楝主要分布区。在广州增城开展了种源苗期试验,试验旨在了解种源间的生长性状地理变异及其规律,初步选择适宜在广东地区造林的苦楝种源,为苦楝造林种子调拨提供试验依据。
1 材料与方法 1.1 材料概况通过查阅苦楝的分布资料[8-9],实地勘察,确定采种点。做到布点均匀,能覆盖整个分布区。采种母树选优标准是:主干明显,干型通直,枝下高在5 m以上,无明显的病虫危害。采种点内母树之间相距远于100 m。各采种点地理位置和海拔见表 1。
|
|
表 1 参试种源概况 Table 1 Summary of different Melia azedarach provenances collected for this study |
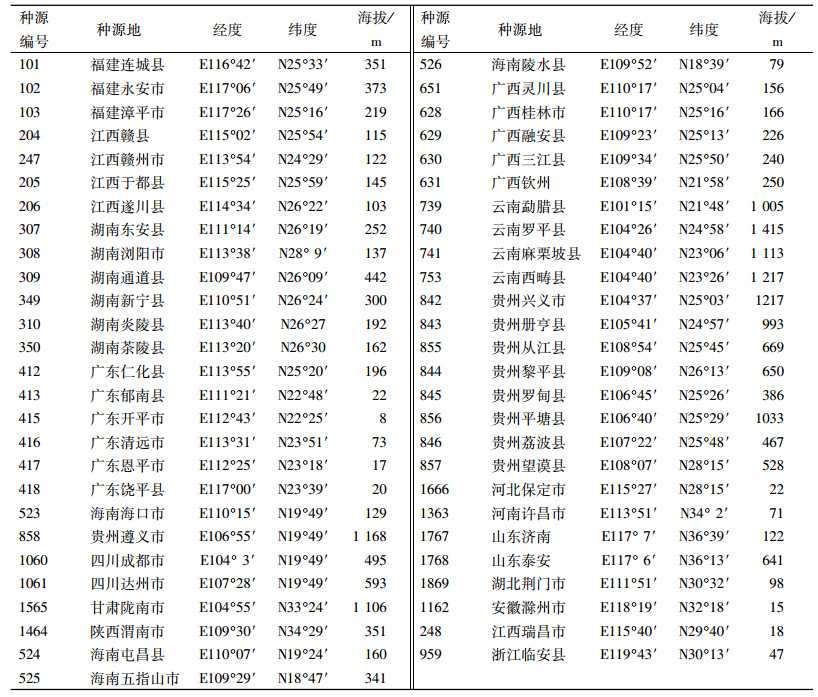
2014年2月,在华南农业大学启林北苗圃进行育苗,当年6月9日移苗造林,参试种源53个(表 1)。造林试验地设在广州增城新塘镇华南农业大学教学科研基地。该地位于东经113°46′、北纬23°20′,属海洋性亚热带季风气候。年降水量为1 931.3 mm,年均气温为21.8 ℃,年均相对湿度为79%,年日照为1 857.9 h。定植株行距为1 m×2 m,植苗穴规格为50 cm×50 cm×50 cm。随机区组设计,7个区组。每小区5株,每个栽植穴施有机肥3 kg。
1.3 试验调查及分析方法 1.3.1 试验设计2015年4月4日对试验幼林进行生长调查。调查性状为树高、地径、侧枝数、干型、东西冠幅、南北冠幅等。用标尺测量树高、东西冠幅、南北冠幅,用游标卡尺测量地径。干型分级如下:1级为有明显主干,且主干通直;2级为有明显主干,主干稍弯曲;3级为有主干,且主干弯曲程度很大;4级为没有主干,多分枝。冠幅起测标准:从种植时30 cm开始,低于30 cm以下且树高相对种植时无变化,表明苗木由于某种原因停止生长,失去试验的目的,记录冠幅为缺失值。
1.3.2 方差分析模型
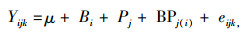
|
式中,Yijk为i区组j种源中的k单株某性状的测量值;μ为试验平均值;Bi为i区组的效应值;Pj为j种源效应值;BPj(i)为i区组与j种源的交互效应值;eijk为残差效应值[10]。
1.3.3 参数计算公式变异系数(CV)[11]:CV=(S/X)×100%,式中, S为标准差,X为平均值。
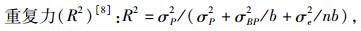
|
式中:R2为种源某性状的重复力,σP2为某性状种源的方差分量,σBP2为区组与种源的互作方差分量,σe2为误差项方差分量,n为小区内株数,b为区组数。
1.3.4 环境因子数据获取气象数据取自各采种点或相近气象台站点在中国气象数据网中国气象资料年值数据集,1951—2012年的气象数据均值。地理因子为采种点GPS记录数据。
采用R和SPSS 19.0统计软件对测量的6个性状进行数据分析。
2 结果与分析 2.1 参试种源间生长量差异分析各种源在6个性状间均存在巨大的差异。种源间在树高性状上的差异幅度为5~280 cm,相差56倍,在地径性状上的差异幅度为1.2~64.0 mm,相差53倍;在干型性状上的差异最小,为1~4,相差3倍;侧枝数的变异幅度为1~18个,东西冠幅和南北冠幅分别为3~200和3~220 cm。经单因素方差分析(表 2),除干型在种源间的差异达显著(P < 0.05)水平外,其余5个性状在种源间的差异均达到极显著(P < 0.01)水平。
|
|
表 2 苦楝种源幼林期生长性状变异分析 Table 2 Analysis of variations in growth traits of young forest of different Melia azedarach provenances |
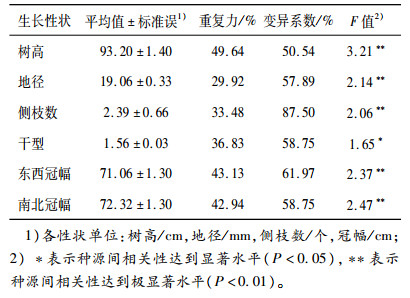
由表 2还可知,树高、东西冠幅和南北冠幅等性状的重复力较高,均达到40%以上,说明这些性状较另外3个性状在种源间的差异更为稳定,而地径重复力仅为29.92%,表明地径在种源间表现的差异稳定性较弱。
为进一步研究区组的立地环境对各性状的影响,进行了种源与区组双因素方差分析。结果(表 3)表明,各个性状在种源间的差异和在区组间的差异均达到极显著水平,表明区组环境对苦楝生长性状的影响也大。除干型外,其他性状的种源与区组的交互效应都达到极显著水平,且方差分量比值比种源间和区组间都高,说明不同种源在不同立地条件下有不同反应。此外,也看出机误的方差分量比值最大,说明种源内个体间的差异以及环境条件造成的差异是很大的。
|
|
表 3 苦楝生长性状的双因素方差分析1) Table 3 Variations in growth traits of Melia azedarach based on double factorial analysis of variance |
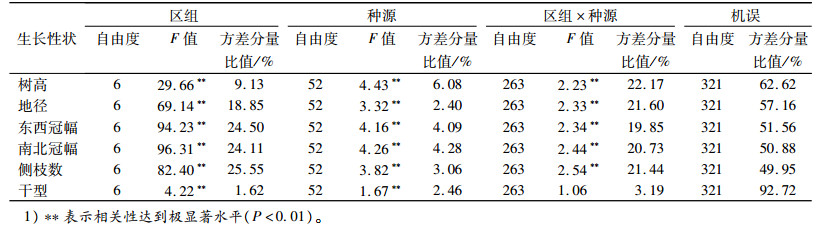
综合生长性状中的树高和地径的多重比较结果(表 4)可知,云南罗平种源(740)生长最快,树高达到135.68 cm以上,地径达到25.41 mm以上,其余3个生长快、生物量大的种源依次为:广西融安种源(629),树高达到131.69 cm以上,地径达到25.75 mm以上;贵州册亨种源(843),其树高达到120.91 cm以上,地径达到25.80 mm以上;湖南新宁种源(349),其树高达到123.72 cm以上,地径达到23.16 mm以上。生长最慢的为山东济南种源(1767),树高在62.50 cm以下,地径在11.50 mm以下。
|
|
表 4 种源间树高与地径差异的多重比较结果1) Table 4 The Duncan analysis of height and diameter comparing different Melia azedarach provenances |
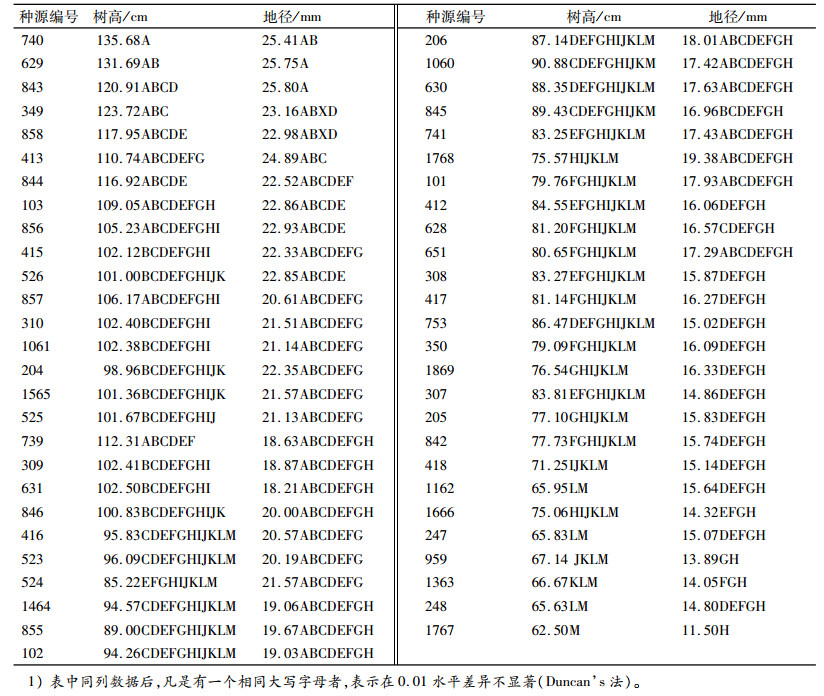
由表 5可知,种源树高与采种点纬度相关性没有达到显著水平,与经度达到显著相关,说明本试验点树高的地理变异的表现以经向变异为主;地径、侧枝数和冠幅与经度的相关性没有达到显著水平,与纬度的相关性达到显著或极显著水平,说明地径、侧枝数和冠幅的地理变异主要受纬度控制;干型与纬度和经度的相关系数均没有达到显著水平。海拔与生长性状的相关性均未达到显著水平。从相关性大小来看,只有树高与纬度的相关系数绝对值较其与经度的相关系数绝对值小,其他各性状与纬度的相关系数绝对值都较其与经度的相关系数绝对值大。
|
|
表 5 生长性状与地理因子的偏相关分析1) Table 5 Partial correlation analysis of growth traits and geographic factors |
综上所述,采种点的纬度与种源的地径、侧枝数、冠幅呈显著或极显著负相关,经度与树高呈显著负相关,表明采种点由南到北,由西到东,苦楝生长变慢,生物量变低。
2.5 各性状与气候因子典型相关性分析对53个种源各生长性状与采种点主要气候因子作典型相关分析[12] (表 6),结果表明:第一典型变量气候因子中,气温、湿度的载荷都在0.5以上,其次是降水量,为0.401。生长性状中,冠幅的载荷最大,都在0.8以上,其次是树高、地径、侧枝数。这说明,气温、湿度对冠幅的生长地理变异影响作用最大,其次是降水量;第二典型变量气候因子中,极端最高本站气压、平均最低气温的载荷都在0.6以上,其次是平均气温、平均最高气温和降水量,均达0.5以上,而生长性状中,树高和地径的载荷为负值,绝对值大于0.5。这说明气压和气温对树高和地径的生长地理变异影响较大,来自极端气压高和平均最低气温低的地区的苦楝,其树高和地径生长较慢;在第三典型变量中,极端最高本站气压、日照百分率和日照时数载荷绝对值均大于0.3,观测性状中干型的载荷大于0.3。这说明极端最高本站气压、日照百分率和日照时数对干型的影响作用最大。
|
|
表 6 生长性状与气候因子的典型相关分析 Table 6 Canonical correlation analysis of growth traits and climatic factors |
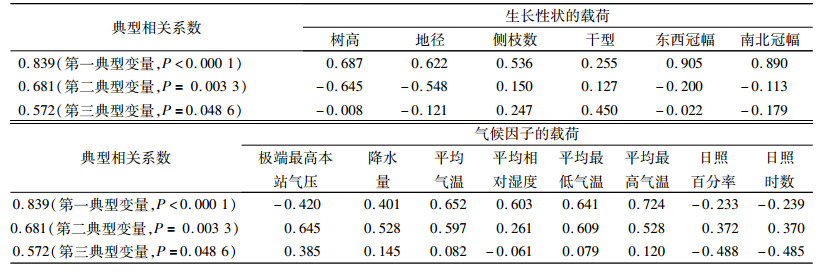
综上所述,来自气温较高、降水丰富、气压低、平均最低气温高、日照丰富地区的种源在广州生长快,生物量大。
2.6 种源聚类分析根据种源树高、地径、侧枝数、干型、冠幅等生长性状,对有效数据进行标准化后,采用欧氏距离离差平方和进行聚类,结果见图 1。结果表明,当欧式距离在1.20时,53个种源可以大致划分为6个类群,第Ⅰ类群为种源101、417、651、418、628、206、1060、307、350、753、412、842、1869、1768,主要来自福建、广东、广西等华南地区;第Ⅱ类群为种源247、1666、1162、1363、248、959、308、1767,主要来自山东、河南、河北、安徽等偏北地区;第Ⅲ类群为种源102、845、309、857、855、205,主要来自贵州、湖南等中部地区;第Ⅳ类群为种源631、103、415、523、525、630、846、204、844、856、416、741、739,主要来自福建、广东、广西、海南等南部沿海地区;第Ⅴ类群为种源524、526、310、1464、1061、1565、349、740、858,主要来自陕西、甘肃、贵州等偏西北地区;第Ⅵ类群包括413、629、843种源,主要来自广东、广西。

|
图 1 53个种源聚类分析结果 Figure 1 Cluster analysis of 53 provenances |
种源试验是林木遗传改良的基础工作之一,探究林木群体变异规律性,并为进一步开展优良种源选择、杂交育种亲本选配提供试验依据。苦楝是我国经济利用价值高,抗逆性强的乡土树种,在各方面性状表现良好,具有广泛的遗传基础。开展苦楝种质资源收集、测定和评价工作,充分利用丰富的苦楝遗传资源,大力挖掘苦楝作为木材或新型生物农药的经济价值,对进行集约种植及产业化开发具有重要现实意义[13]。
各种源在6个性状间均存在巨大的差异,其中,树高和地径在种源间的差异最大。经单因素方差分析,除干型在种源间的差异达显著(P < 0.05)水平外,其余5个性状在种源间的差异均达到极显著(P < 0.01)水平。树高、东西冠幅和南北冠幅等性状的重复力较高,达到40%以上,说明这些性状较其他3个性状在种源间的差异更稳定,而地径重复力仅为29.92%。
苦楝种源的地径、侧枝数、冠幅与采种点的纬度呈显著或极显著负相关,且树高与经度呈显著负相关,表明采种点由南到北,由西到东,苦楝生长变慢,生物量降低。陈丽君等[7]对苦楝70个种源的果核和种子的18个性状进行了观测和分析,发现苦楝种子由南到北存在明显地理变异。程诗明[8]将24个苦楝群体729个家系的5个地理气候因子及与其对应的苦楝18个叶片、种子等表型性状因子进行典型相关分析,研究得出苦楝群体表型性状受经度、纬度的双重控制, 随着纬度、经度梯度变异明显, 具很强的东北、西南变异趋势[14]。本研究所选取的苦楝种源更为丰富,各生长性状也多以纬度变异为主,但树高在本试验中呈经度变异为主,还需进一步试验观察。
苗木生长性状的地理变异具有明显的气候生态基础,受水分、气温、湿度、气压的多重影响。根据典型相关分析,由第一典型变量可知,采种点气温、湿度对冠幅、树高和地径生长的地理变异影响作用最大,其次是降水量;由第二典型变量可知,极端最高本站气压和平均最低气温对树高和地径生长地理变异影响最大,其次是降水量;由第三典型变量可知,极端最高本站气压、日照百分率和日照时数对干型的影响作用最大。总体上看,来自气温较高、降水丰富、气压低、平均最低气温高、日照丰富地区的种源在广州生长快,生物量大。
根据种源幼林生长性状采用欧氏距离离差平方和进行聚类,将53个种源可以大致划分为6个类群,分类结果的地理格局较为明显。陈丽君等[7]的分析结果表明,根据果核和种子的性状对苦楝种源聚类,广东、广西多为一类,贵州、甘肃也多为一类,这与本研究结果有一致性。程诗明等[9, 15]将中国苦楝分布区划分为11个物候区,将本研究的聚类结果与苦楝物候区进行对照,相同区域的种源基本上可归属在同一个气候区。
| [1] |
马万侠, 陈民生, 宋尚文, 等. 苦楝半同胞家系苗期生长性状的研究[J]. 山东农业大学学报(自然科学版), 2010, 41(1): 27-30. DOI:10.3969/j.issn.1000-2324.2010.01.006 (  0) 0) |
| [2] |
林盛松, 李小莲, 肖伏德, 等. 寿宁县不同海拔高度苦楝生长效应分析[J]. 福建林业科技, 2011, 38(4): 41-45. DOI:10.3969/j.issn.1002-7351.2011.04.10 ( 0) |
| [3] |
董玉峰, 姜岳忠, 马玲, 等. 苦楝遗传改良研究进展[J]. 山东林业科技, 2010, 40(4): 104-107. DOI:10.3969/j.issn.1002-2724.2010.04.031 ( 0) |
| [4] |
廖柏勇, 陈晓阳, 陈丽君, 等. 苦楝种源间种子发芽变异的观测[J]. 广东农业科学, 2014, 41(11): 43-47. DOI:10.3969/j.issn.1004-874X.2014.11.010 ( 0) |
| [5] |
教忠意, 徐长柏, 丁学农. 苦楝优良家系初步选择[J]. 浙江林业科技, 2011, 31(3): 48-50. DOI:10.3969/j.issn.1001-3776.2011.03.012 ( 0) |
| [6] |
陈羡德, 陈礼光, 阙茂文, 等. 不同种源苦楝幼林试验初步研究[J]. 福建林学院学报, 2008, 28(1): 23-26. DOI:10.3969/j.issn.1001-389X.2008.01.006 ( 0) |
| [7] |
陈丽君, 邓小梅, 丁美美, 等. 苦楝种源果核及种子性状地理变异的研究[J]. 北京林业大学学报, 2014, 36(1): 15-20. ( 0) |
| [8] |
程诗明. 苦楝聚合群体遗传多样性研究与核心种质构建[D]. 北京: 中国林业科学研究院, 2005. http://cdmd.cnki.com.cn/article/cdmd-82201-2005128807.htm
( 0) |
| [9] |
程诗明, 顾万春. 苦楝中国分布区的物候区划[J]. 林业科学, 2005, 41(3): 186-191. DOI:10.3321/j.issn:1001-7488.2005.03.032 ( 0) |
| [10] |
黄少伟, 谢维辉. 实用SAS编程与林业试验数据分析[M]. 广州: 华南理工大学出版社, 2001, 246.
( 0) |
| [11] |
李宗艳, 李名扬. 硬叶兜兰居群表型变异研究[J]. 植物遗传资源学报, 2015, 16(4): 765-771. ( 0) |
| [12] |
陈晓阳. 典型相关分析在树种地理变异研究中的应用[J]. 北京林业大学学报, 1990. ( 0) |
| [13] |
陈晓阳, 沈熙环. 林木育种学[M]. 北京: 高等教育出版社, 2005, 51-52.
( 0) |
| [14] |
程诗明, 顾万春. 苦楝中国分布区的物候区划[J]. 林业科学, 2005, 41(3): 186-191. DOI:10.3321/j.issn:1001-7488.2005.03.032 ( 0) |
| [15] |
程诗明, 顾万春. 苦楝表型区划的研究[J]. 林业科学研究, 2006, 19(3): 337-341. DOI:10.3321/j.issn:1001-1498.2006.03.014 ( 0) |