 2016, Vol. 37
2016, Vol. 37


2. 河南科技大学 林学院,河南 洛阳 473001
2. College of Forest, Henan University of Science and Technology, Luoyang 473001, China
鹞落坪半夏Pinella yaoluopingensis属天南星科半夏属植物,为安徽省鹞落坪国家自然保护区特有种[1],分布范围狭窄。鹞落坪半夏与半夏P. ternata从外形看较为相似,但鹞落坪半夏叶柄无珠芽,而半夏叶柄有珠芽,这是两者最主要的外形区别[2-3]。半夏在民间用于止咳已历史悠久,鹞落坪半夏同样具有明显的止咳作用,止咳强度与半夏近似,但作用要快于半夏[4],是优良的药材资源。
生态位理论是当代生态学极受关注的理论之一,也是种群生态研究的核心问题[5],被广泛用于研究物种共存机制[6-7],它是评价种间和种内关系及种群在群落中地位的重要手段[8]。刻画生态位特征的参数常用指标主要有生态位宽度和生态位重叠,生态位宽度反映种群对环境适应的状况或对资源利用的程度[9-10],生态位重叠反映2个或多个种群在适应环境和利用资源的实际幅度或潜在能力方面所表现出的共同性或相似性[9]。生态位现已被广泛应用于植物资源的保护与利用、生物多样性保护及群落演替等研究[11-12]。鹞落坪半夏分布范围狭窄,主要分布在黄山松Pinus taiwanensis疏林下,林下种植有天麻Gastrodia elata,或为弃耕地,鹞落坪半夏仅以伴生种出现在群落中,是狭域分布的特有种,数量有限。本文通过对鹞落坪半夏生境地主要草本种群的生态位特征进行分析,探讨各种群对环境资源的利用状况以及物种之间的关系,以期为制定鹞落坪半夏保护措施及对鹞落坪半夏资源开发利用提供指导性建议。
1 研究地自然概况鹞落坪国家级自然保护区地处皖鄂2省3县(安徽岳西、霍山和湖北英山)交界处的安徽安庆市岳西县包家乡境内,其西接湖北英山,北邻大别山主峰白马尖,地理坐标为N30°40′~31°06′,E116°03′~116°33′,总面积123 km2。保护区内峰峦林立,溪谷众多,整个地势向东北缓降。其主峰多枝尖海拔1 721.5 m,为大别山第3高峰,最低处鲤鱼尾海拔500 m。该区属北亚热带季风气候区,空气湿润,气温较低,雨雪充沛,年降水量1 400~2 000 mm,年平均温度15 ℃,无霜期210~220 d。本区土壤类型有山地棕壤、山地黄棕壤、山区草甸土和水稻土,山地棕壤为垂直带谱土壤。该保护区植物区系起源古老,特有现象明显,植物种类丰富,植被类型多样,地带性植被属于北亚热带常绿落叶阔叶混交林,但常绿阔叶树种比重不大,主要植被为优势种不明显的大范围的落叶阔叶林[13-15]。
2 研究方法 2.1 野外调查在经过2012—2014年多次踏查和走访的基础上,于2014年4月下旬在鹞落坪保护区鹞落坪半夏集中分布的大湾、王湾、人参场及保护区管理处后方山坡,采用典型取样法,在鹞落坪半夏生境地设置4块样地进行调查(每块样地400 m2)。在每一样地的4角分别设置4个1 m×1 m的小样方,记录每个小样方内的草本植物种类、多度、盖度、高度等指标。记录样地海拔、坡度、坡向、干扰情况等信息。
2.2 数据处理 2.2.1 重要值计测用种群的重要值作为指标进行生态位分析。先用公式计算每个小样方各种群重要值(Ⅳ):Ⅳ =(相对盖度+相对密度+相对高度)/3,然后对所有小样方的重要值进行合计取平均值,获得鹞落坪半夏生境地各草本种群的重要值。重要值越大表明种群在群落中的优势度越高,剔除仅出现1次的偶见种,选取重要值较大的草本种群作为主要种群进行后续研究。
2.2.2 生态位计测重要值作为物种的综合数量指标,表征物种在群落中的地位和作用,是应用最广的物种特征值[16],以重要值为指标测定群落中种群的生态位宽度和生态位重叠,能较好地表达群落中种群生态位的对比关系。为此,本文以样地作为资源状态,以物种重要值为指标分析鹞落坪半夏生境地主要草本种群的生态位宽度和生态位重叠。
生态位宽度的计算:采用Levins和Shaanon-Wiener计算方式[10, 17-19]。
Levins生态位宽度(Bi)指数公式为:
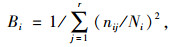
|
(1) |
Shaanon-Wiener生态位宽度(B′i)指数公式为:
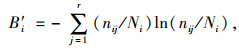
|
(2) |
式中,nij为种群i利用资源状态j的数量(本文以种群i在第j样方的重要值表示)。Ni为种群i的总数量,r为样方数。
生态位重叠的计算:采用Levins生态位重叠指数[16, 19]。
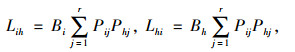
|
(3) |
式中,Lih、Lhi分别为物种i对物种h、物种h对物种i的生态位重叠指数,Bi、Bh分别为物种i、h的生态位宽度,Pij、Phj分别为种群i和种群h在第j个样方重要值占其重要值总数的比例。
2.2.3 相关系数计测采用SPSS软件(13.0)计算各种群重要值(Ⅳ)与生态位宽度(Bi和Bi′)之间以及生态位宽度Bi和Bi′之间的相关性。
3 结果与分析 3.1 主要草本种群重要值本研究共调查4个样地16个样方,记录到41个草本种群。重要值居前3位的依次是博落回Macleaya cordata、金星蕨Parathelypteris glanduligera、臭草Melica scabrosa,它们的重要值在12~15之间,其他草本种类的重要值均在6以下,鹞落坪半夏重要值位居第11位,为2.673。剔除偶见种后,选取重要值较大的17个种群进行生态位分析,该17个种群重要值见表 1。
|
|
表 1 鹞落坪半夏生境地主要草本种群重要值和生态位宽度 Table 1 The important values and niche breadths of main herbaceous populations in habitat of Pinella yaoluopingensis |
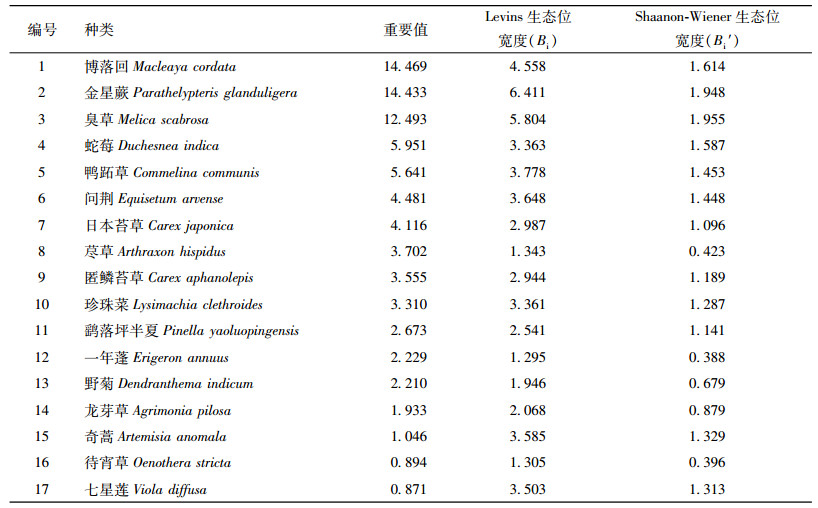
鹞落坪半夏生境地17个主要草本种群生态位宽度值见表 1。结合生态位宽度Bi和Bi′来看,鹞落坪半夏生境地主要草本种群生态位宽度居前3位的种群是金星蕨、臭草、博落回,该3个种群的Bi分别是6.411、5.804、4.558,对应的Bi′分别是1.948、1.955、1.614。该3个种群的重要值位居群落草本层的前3位,是鹞落坪半夏生境地草本层组成的优势种群。鹞落坪半夏Bi和Bi′值分别为2.541、1.141,分别位于第12、11位,其重要值位居第11位。生态位宽度居最后3位的是荩草Arthraxon hispidus、待宵草Oenothera stricta、一年蓬Erigeron annuus,其中只有荩草重要值居中等,其他种类的重要值均在第11位以后。鹞落坪半夏生境地主要草本种群生态位宽度分布格局见表 2。从表 2可以看出,多数种群的Bi集中于1.0~4.0,占所有种群的82.35%,其中又多集中于3.0~4.0,占所有种群的35.29%;多数种群的Bi′集中于0.3~1.5,占76.67%,其中又多集中于1.2~1.5,占29.41%。可见,多数种群的生态位宽度较大。
|
|
表 2 鹞落坪半夏生境地主要草本种群生态位宽度分布格局 Table 2 The niche breadth distribution patterns of main herbaceous populations in habitat of Pinella yaoluopingensis |
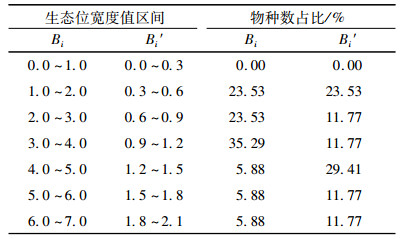
博落回、金星蕨、臭草有较大的生态位宽度,与它们分布范围广、数量较多有关,博落回在3个样地6个小样方有分布,金星蕨在3个样地8个小样方有分布,臭草在4个样地9个小样方有分布,它们对环境适应能力及对各种资源利用能力和竞争能力较强。而荩草、待宵草、一年蓬生态位宽度较低,与它们在鹞落坪半夏生境地出现的频度低有关,该3个种群均仅分别在1个样地2个小样方出现,对资源的利用程度相对较低。本研究采用的是典型取样法,针对有鹞落坪半夏集中分布的区域设置群落样地,但鹞落坪半夏生态位宽度并未有优势,这与鹞落坪半夏在生境地空间分布不均匀、数量不多是相吻合的。鹞落坪半夏在4个样地16个小样方中仅有4个小样方有分布,而对于生境地的金星蕨、臭草可高达8~9个小样方有分布,可见,适宜鹞落坪半夏的生境少,其对所处的环境条件比较苛刻。本研究以重要值作为资源利用的参数,由其计测得出的生态位宽度具有多维意义[20]。从光照条件来看,有鹞落坪半夏的4个小样方的乔木层郁闭度均在0.4~0.6之间,而对于郁闭度较低或较高的小样方则无鹞落坪半夏的出现。另外,从踏查的结果来看,在密林中或田间地头也未发现有鹞落坪半夏的分布。可见,群落光照条件可能是影响鹞落坪半夏分布的一个因素。土壤条件等在空间的分布往往是异质性的,有不少学者已证实植物生态位分化的环境资源维度包括土壤质地、土壤养分、土壤水分等方面[21]。因此,除了光照条件的影响,土壤条件等可能也是影响鹞落坪半夏生态位宽度相对不宽的因素。
针对上述结果中种群生态位宽度与重要值开展相关性分析,结果表明,17个种群的重要值Ⅳ与生态位宽度Bi相关系数为0.807(Sig.=0.000),Ⅳ与Bi′相关系数为0.707(Sig.=0.002)。可见,17个种群的重要值与生态位宽度呈极显著正相关。另外,17个主要种群的Bi和Bi′相关性呈极显著,相关系数为0.952(Sig.=0.000),这说明了17个主要种群生态位宽度Bi和Bi′排序是比较一致的,由此也认为,本研究计测得到的17个主要草本种群生态位宽度Bi和Bi′是比较可靠的。
在鹞落坪半夏生境地中,17个主要草本种群重要值排序与生态位宽度排序具有一致性,但也并非完全一致。如七星莲Viola diffusa,其Bi居第7位,Bi′居第8位,该种群在3个样地中4个小样方有分布,分布范围较广,但其重要值却是最低的,主要原因是七星莲个体矮小以致于其重要值较低。又如荩草,其生态位Bi和Bi′均为第15位,但其重要值却居第8位,这是因为荩草仅在2个小样方中出现,虽然出现频度低,但在1个小样方中的荩草的覆盖度却较有优势。以上说明了重要值和生态位宽度2种指标各有侧重,重要值主要表示种群在群落中的优势程度,而生态位宽度则主要体现种群对资源的利用能力。
3.2.2 生态位重叠鹞落坪半夏生境地主要草本种群间生态位重叠值见表 3。经统计,17个主要草本种群形成的272个种对中(包括Lhi和LihS),生态位重叠值>1.0的种对仅有3对,仅占总物种对的1.10%,这3个种对分别为鹞落坪半夏-荩草、蛇莓Duchesnea indica-龙芽草Agrimonia pilosa、待宵草-一年蓬;生态位重叠值在0.8~1.0的也是3对,为一年蓬-待宵草、匿鳞苔草Carex aphanolepis-野菊Dendranthema indicum、金星蕨-野菊;生态位重叠值在0.6~0.8之间的有9对,占3.31%,为珍珠菜Lysimachia clethroides-野菊、博落回-鸭跖草Commelina communis、金星蕨-奇蒿Artemisia anomala、问荆Equisetum arvense-待宵草、龙芽草-蛇莓、鸭跖草-博落回、问荆-一年蓬、臭草-荩草、荩草-鹞落坪半夏;生态位重叠值在0.4~0.6之间的有30对,占所有种对的11.03%。综上所述,生态位重叠值在0.4以上的种对仅45对,仅占总对数的16.54%。生态位重叠值在0.4以下的种对占所有种对的83.46%,其中生态位重叠值为0的种对占38.60%,这表明这些种对生态学特性相似不大,对资源的共享趋势不明显,这一研究结果与鹞落坪半夏生境地种间联结研究结果相吻合(另文发表)。
|
|
表 3 鹞落坪半夏生境地主要草本种群生态位重叠1) Table 3 The niche overlaps of main herbaceous populations in habitat of Pinella yaoluopingensis |
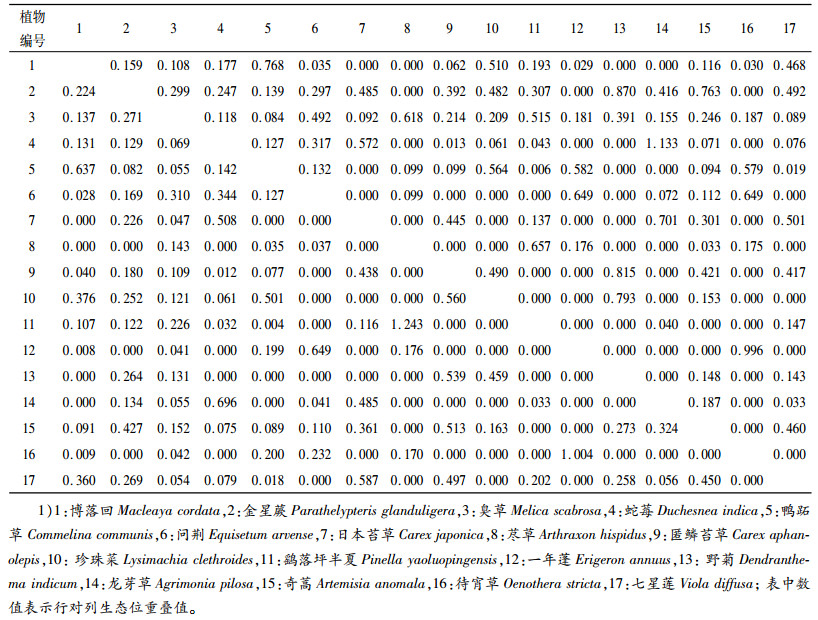
生态位重叠值高的种对间对生境的要求相对一致,经常相伴出现[16]。较高的生态位重叠意味着种群之间对于资源环境具有相似的生态学要求[22]。在这些重叠值较高的15个种对(生态位重叠值>0.6)中,种群生态位宽度均比较大的仅有2个种对,即博落回-鸭跖草、鸭跖草-博落回,博落回Bi和Bi′均排列第3位,鸭跖草Bi和Bi′分别排列第4、5位。生态位重叠值较大的15个种对中,生态位宽的种群与生态位窄的种群有5对较大重叠值的种对,比例较大,占1/3,如臭草-荩草、金星蕨-野菊等种对,臭草的Bi和Bi′排列第2和第1位,荩草的Bi和Bi′均排列第15位,金星蕨的Bi和Bi′排列第1和第2位,野菊的Bi和Bi′均排列第14位。2个种群生态位均窄但生态位重叠高的种对数量也较多,有3对。最典型的是鹞落坪半夏-荩草这一种对,这个种对的生态位重叠值在所有种对中最高,达1.243,而鹞落坪半夏的Bi和Bi′分别为第12和第11位,荩草的Bi和Bi′均居第15位。可见,生态位重叠值高的种对不都由生态位宽的种群构成。
结合表 1和表 3可以看到,生态位宽度居前3位的为金星蕨、臭草、博落回,它们之间的生态位重叠值并不高,均在0.3以下,与这些种群本身的生态学特性不尽相同对资源的要求不一致有关。影响植物生态学特性因素有气候因子和土壤条件等,本研究所设置的4个样地地理位置相差不远,气候因子温度、水分等对3个种群的分布影响不大,另外,从样方调查结果来看,3个种群的分布也并未明显受坡向影响,那么鹞落坪半夏生境地主要草本种群空间上的分布可能主要与土壤条件及群落光照条件异质性有关。博落回与金星蕨、臭草对资源的不同要求,从它们所在环境的光照条件可以得到体现,分布有博落回的环境总体上乔木层的郁闭度较低或灌木层的盖度较小,而金星蕨、臭草所在的环境总体上乔木层的郁闭度较高或灌木层的盖度较大。金星蕨、臭草所在的环境上层郁闭度或盖度相似,但两者生态位重叠值并不高,那么金星蕨、臭草对土壤条件如土壤质地、土壤养分、土壤水分等的要求不同可能是影响两者空间分布不同的主要因素。有不少种群与鹞落坪半夏之间生态位重叠值为0,这也说明它们之间的生态学特性相差甚远,由于各自需要的小环境有所不同,导致它们不出现在一起。
进一步分析发现,Lih和Lhi相比,Lih相对较大,生态位重叠值大于0.6的Lih有11对,而Lhi仅有4对。出现这种前一个种群对后一个种群的重叠值较高而后一个种群对前一个种群的重叠值较低的现象,主要是由于它们分别是由各自种群的生态位宽度指标确定的[16, 23],如鹞落坪半夏、荩草Bi分别为2.541、1.343,鹞落坪半夏-荩草生态位重叠值为1.243,而荩草-鹞落坪半夏生态位重叠值为0.657,说明生态位较宽的种群对生态位较窄的种群可能有较高的重叠值。
总的来看,鹞落坪半夏生境地主要草本种群生态位重叠值不高,多数种群之间生态学特性相似性不大,彼此之间能和谐生长在同一群落。同时可见,鹞落坪半夏生境地主要草本种群的生态位宽度和生态位重叠之间并不存在直接的线性关系。
4 讨论生态位宽度主要通过种群对资源的利用能力来评价某一种群在群落中的地位和作用[23-25],生态位宽度作为种群对环境资源利用多样性的一种测度,决定于该种群的形态适应、生理反应和特有的行为[26],种群的生态位宽度越大,说明该种群在群落中的地位越高,其分布的范围越广,对资源的利用能力越强[27]。鹞落坪半夏生境地主要草本种群中,金星蕨、臭草、博落回、鸭跖草等生态位宽度较宽,表明它们在鹞落坪半夏生境地内分布较广、数量较多、利用资源较为充分,具有较强的生态适应性,生存机会较多。而一年蓬、待宵草、荩草等生态位宽度较窄,有些种群虽然在个别样地优势明显,但由于它们在鹞落坪半夏生境地出现的频度较低,因此所利用的资源能力低。生态位宽度能很好地反映种群的生态适应性和利用资源的能力,而重要值是综合衡量物种在群落中地位和作用的有效指标[28],在鹞落坪半夏生境地中,主要草本种群重要值排序与生态位宽度排序呈极显著正相关,但也有少数一些种群的生态位宽度和重要值排序不一致,这说明了2种指标各有侧重,重要值主要表示种群在群落中的优势程度,而生态位宽度则主要体现种群对资源的利用能力。一些学者认为,不同种群在一定区域下会选择不同的生存对策,如重要值大生态位也宽的种群,因对环境适应能力强,是通过扩大地理分布区来维持足够的种群数量,而重要值大生态位窄的种群,会选择在适宜的环境中保持优势数量来维持足够的种群数量[23, 29]。
生态位宽度和生态位重叠是2种重要的生态位特征,生态位宽度可以反映种群对资源的利用程度,生态位重叠体现种群对同等级资源的利用程度以及配置关系[16]。本研究结果显示生态位宽的2个种群重叠值并不一定高,生态位窄的2个种群的重叠值也并不一定低,这一结果与其他群落种群之间生态位宽度与生态位重叠研究结果是一致的[20, 25-26, 30-33]。刘帅华等[28]对青海大通不同人工林林下草本群落生态位研究时,总结出较高生态位重叠的出现总体上有3种情况:①生态位较宽的2个种群生态位重叠值较大;②具有较宽生态位和具有较窄生态位的2个种群间存在较大的生态位重叠;③生态位较窄的2个种群间具有较大的生态位重叠。本研究结果与上述结果相一致,但总的来说,生态位较宽的种群与其他种群生态位重叠的机会要大一些,而生态位较窄的种群与其他种群生态位重叠的机会要小些,这与不少学者的研究结果是一致的[21, 32-36]。究其原因,可能是生态位宽度较大的种群对资源的利用能力较强,分布广泛,因而可能会与其他种群有较大的重叠。生态位较窄的2个种群之间也可能具有大的生态位重叠值,贺山峰等[25, 37-38]认为,生态位宽度均窄的种群而生态位重叠值大的原因可能是2个种群之间或有着很相似的生态学习性,或对环境因子的要求具有互补性,2个种群出现的几率不高、数量不大,但是两者却经常出现在一起,而在适宜生境斑块以外的空间种群的分布极为贫乏。
半夏生长的环境较为多样,可生长于草坡、荒地、玉米地、田边或疏林下[39],而鹞落坪半夏生长环境较苛刻。从笔者对鹞落坪半夏的生态环境调查来看,鹞落坪半夏在鹞落坪保护区主要分布在黄山松疏林下。徐如松等[40]在研究旅游干扰对鹞落坪保护区植被影响中也提到鹞落坪半夏是对环境依赖性强的物种,喜湿耐荫。因此,要保护好鹞落坪半夏,其所处环境的上层林木应保护好。目前调查发现鹞落坪半夏生境地离现有旅游线路并不远,有旅游就必然存在自然保护和旅游开发的矛盾,笔者认为,发展生态旅游应尽量减少旅游对环境的干扰,特别是对珍稀濒危植物及特有植物所在环境的干扰。对环境破坏最具影响力的是对群落乔木层的破坏,值得欣慰的是目前黄山松尚保护较好,生态旅游对鹞落坪半夏并未造成威胁。群落内其他种类对鹞落坪半夏的影响会如何呢?从鹞落坪半夏生境地17个主要草本种群生态位重叠值为0的种对数(105对)和重叠值在0.4以下种对所占的比例(83.46%)来看,鹞落坪半夏生境地主要草本种群能和谐生长在同一群落,在鹞落坪半夏与其他种群构成的种对中,只有鹞落坪半夏-荩草之间的重叠值大于0.6。由鹞落坪半夏与荩草生态位宽度相比较,鹞落坪半夏具有较大的生态位宽度,荩草并未对鹞落坪半夏构成威胁。因此,从目前来看,鹞落坪半夏总体上能与其生境地主要草本种群和谐地生长于群落中。但从生态位宽度排序来看,鹞落坪半夏的地位并没有优势,在群落中仅处于伴生的地位,占优势的是金星蕨、臭草、博落回。如果群落无人为干扰,随着群落不断演替发展,乔木层郁闭度越来越高,更高的郁闭度有可能对草本层优势种金星蕨、臭草比较有利,这些草本层优势种的优势地位有可能越来越明显,而作为群落伴生种的鹞落坪半夏是否能竞争得过这些种类而与群落中的其他种类稳定地共存呢?尚需对群落的发展演替给予更多的关注,以更好地保护该狭域特有种及对资源进行有效开发利用。
| [1] |
谢中稳, 蔡永立. 安徽省鹞落坪自然保护区植物区系基本特征的研究[J]. 安徽农业大学学报, 1994, 21(4): 507-512. (  0) 0) |
| [2] |
刘晓龙, 郭新弧. 安徽半夏属一新种[J]. 云南植物研究, 1986, 8(2): 223-224. ( 0) |
| [3] |
顾德兴, 李云香, 徐炳声. 半夏的繁殖生物学研究[J]. 植物资源与环境学报, 1994, 3(4): 44-48. ( 0) |
| [4] |
桂常青, 黄志力, 伍必英, 等. 鹞落坪半夏的药理作用研究[J]. 中药药理与临床, 1998, 14(4): 27-28. ( 0) |
| [5] |
LEIBOLD M A. The niche concept revisited: Mechanistic models and community context[J]. Ecology, 1995, 76(5): 1371-1382. DOI:10.2307/1938141 ( 0) |
| [6] |
WRIGHT S J. Plant diversity in tropical forests: A review of mechanisms of species coexistence[J]. Oecologia, 2002, 130(1): 1-14. DOI:10.1007/s004420100809 ( 0) |
| [7] |
THUILLER W, ALBERT C, ARAUJO M B, et al. Predicting global change impacts on plant species' distributions: Future challenges[J]. Perspect Plant Ecol, 2008, 9(3/4): 137-152. ( 0) |
| [8] |
林大影, 邢韶华, 赵勃, 等. 北京山区森林植被中珍稀濒危草本植物生态位[J]. 生态学杂志, 2007, 26(6): 781-786. DOI:10.3321/j.issn:1000-4890.2007.06.002 ( 0) |
| [9] |
WEIDER L J. Niche breadth and life history variation in a hybrid daphnia complex[J]. Ecology, 1993, 74(3): 935-943. DOI:10.2307/1940817 ( 0) |
| [10] |
李德志, 石强, 藏润国, 等. 物种或种群生态位宽度与生态位重叠的计测模型[J]. 林业科学, 2006, 42(7): 95-103. ( 0) |
| [11] |
TILMAN D. Causes, consequences and ethics of biodiversity[J]. Nature, 2000, 405(6783): 208-211. DOI:10.1038/35012217 ( 0) |
| [12] |
JORGE S A, TOWNSEND P. Interpretation of models of fundamental ecological niches and species'distributional areas[J]. Biodiv Inform, 2005, 2: 1-10. ( 0) |
| [13] |
谢中稳, 吴国芳. 安徽省鹞落坪自然保护区的植被类型及其分布[J]. 华东师范大学学报(自然科学版), 1995(3): 93-102. ( 0) |
| [14] |
高林, 徐如松, 曹姣仙, 等. 鹞落坪国家级自然保护区植被类型的初步探讨[J]. 生物学杂志, 2002, 19(6): 32-34. DOI:10.3969/j.issn.2095-1736.2002.06.013 ( 0) |
| [15] |
李珍. 安徽鹞落坪自然保护区维管植物多样性研究[D]. 合肥: 安徽大学, 2007. http://cdmd.cnki.com.cn/Article/CDMD-10357-2007193594.htm
( 0) |
| [16] |
柳江, 洪伟, 吴承祯, 等. 退化红壤区植被恢复过程中灌木层主要种群的生态位特征[J]. 植物资源与环境学报, 2002, 11(2): 11-16. DOI:10.3969/j.issn.1674-7895.2002.02.003 ( 0) |
| [17] |
张金屯. 植被数量生态学[M]. 北京: 中国科学技术出版社, 1995.
( 0) |
| [18] |
王琳, 张金屯. 濒危植物矮牡丹的生态位研究[J]. 生态学杂志, 2001, 20(4): 65-69. DOI:10.3321/j.issn:1000-4890.2001.04.017 ( 0) |
| [19] |
潘丽芹, 季华, 傅强, 等. 濒危植物荷叶铁线蕨的生态群落调查及生态位分析[J]. 扬州大学学报(农业与生命科学版), 2005, 26(4): 74-78. ( 0) |
| [20] |
陈玉凯, 杨琦, 莫燕妮, 等. 海南岛霸王岭国家重点保护植物的生态位研究[J]. 植物生态学报, 2014, 38(6): 576-584. ( 0) |
| [21] |
庞春花, 范晓, 张峰, 等. 不同资源维度上汾河流域下游优势种的生态位[J]. 生态学杂志, 2015, 34(2): 380-386. ( 0) |
| [22] |
SILVERTOWN J W. The distribution of plants in limestone pavements: Tests of species interaction and niche separation against null hypotheses[J]. J Ecol, 1983, 71(3): 819-828. DOI:10.2307/2259595 ( 0) |
| [23] |
王伟伟, 杨海龙, 贺康宁, 等. 祁连山青海云杉人工林草本层生态位及生态响应研究[J]. 草地学报, 2012, 20(4): 626-630. ( 0) |
| [24] |
徐德静, 王鹏鹏, 何跃军, 等. 黔北丹霞地貌桫椤群落优势种群生态位研究[J]. 植物研究, 2014, 34(5): 612-618. ( 0) |
| [25] |
宋晓伟. 小陇山国家级自然保护区森林群落多样性与生态位研究[D]. 兰州: 西北师范大学, 2013. http://cdmd.cnki.com.cn/Article/CDMD-10736-1014019207.htm
( 0) |
| [26] |
霍红, 冯起, 苏永红, 等. 额济纳绿洲植物群落种间关系和生态位研究[J]. 中国沙漠, 2013, 33(4): 1027-1033. ( 0) |
| [27] |
曹培健, 于明坚, 金孝锋, 等. 水盾草入侵沉水植物群落主要种群生态位和种间联结研究[J]. 浙江大学学报(农业与生命科学版), 2006, 32(3): 334-340. DOI:10.3321/j.issn:1008-9209.2006.03.018 ( 0) |
| [28] |
刘帅华, 贺康宁, 董梅, 等. 青海大通不同人工林林下草本群落生态位特征[J]. 四川农业大学学报, 2012, 30(2): 167-173. DOI:10.3969/j.issn.1000-2650.2012.02.009 ( 0) |
| [29] |
魏文超, 何友均, 邹大林, 等. 澜沧江上游森林珍稀草本植物生态位研究[J]. 北京林业大学学报, 2004, 26(3): 7-12. DOI:10.3321/j.issn:1000-1522.2004.03.002 ( 0) |
| [30] |
王立龙, 王广林, 黄永杰, 等. 黄山濒危植物小花木兰生态位与年龄结构研究[J]. 生态学报, 2006, 26(6): 1862-1871. DOI:10.3321/j.issn:1000-0933.2006.06.030 ( 0) |
| [31] |
胡正华, 钱海源, 于明坚. 古田山国家级自然保护区甜槠林优势种群生态位[J]. 生态学报, 2009, 29(7): 3670-3677. DOI:10.3321/j.issn:1000-0933.2009.07.027 ( 0) |
| [32] |
赵发珠, 韩新辉, 杨改河, 等. 陕北典型退耕还林区林下草本物种生态位研究[J]. 西北植物学报, 2011, 31(10): 2090-2099. ( 0) |
| [33] |
唐愽言, 操国兴, 蒋万平. 武陵山大木山自然保护区红豆杉群落优势乔木种群生态位和种间联结[J]. 植物分类与资源学报, 2014, 36(5): 668-674. ( 0) |
| [34] |
郭全邦, 刘玉成, 李旭光. 缙云山森林次生演替序列优势种群的生态位[J]. 西南师范大学学报(自然科学版), 1997, 22(1): 73-78. ( 0) |
| [35] |
WALKER B. Conserving biological diversity through ecosystem resilience[J]. Conserv Biol, 1995, 9(4): 747-752. DOI:10.1046/j.1523-1739.1995.09040747.x ( 0) |
| [36] |
王伟伟, 杨海龙, 贺康宁, 等. 青海高寒区不同人工林配置下草本群落生态位研究[J]. 水土保持研究, 2012, 19(3): 156-160. ( 0) |
| [37] |
贺山峰, 蒋德明, 李晓兰, 等. 小叶锦鸡儿固沙群落草本种群重要值与生态位的研究[J]. 干旱区资源与环境, 2007, 21(10): 150-155. DOI:10.3969/j.issn.1003-7578.2007.10.029 ( 0) |
| [38] |
杨宗慧, 郎学东, 李帅锋, 等. 篦子三尖杉群落优势种群生态位和种间关系[J]. 林业科学研究, 2015, 28(4): 473-478. DOI:10.3969/j.issn.1001-1498.2015.04.004 ( 0) |
| [39] |
中国科学院中国植物志编辑委员会. 中国植物志:第13卷:第2分册[M]. 北京: 科学出版社, 1979, 203-204.
( 0) |
| [40] |
徐如松. 旅游干扰对鹞落坪国家自然保护区植被影响的初步研究[D]. 合肥: 安徽大学, 2009. http://www.wanfangdata.com.cn/details/detail.do?_type=degree&id=D203830
( 0) |