 2016, Vol. 37
2016, Vol. 37



2. 华南农业大学 林学与风景园林学院/广东省森林植物种质资源创新与利用重点实验室,广东 广州 510642;
3. 佛山市林业科学研究所,广东 广州 528222;
4. 广东生态工程职业学院,广东 广州 510520
2. College of Forestry and Landscape Architectural, South China Agricultural University/Guangdong Key Laboratory for Innovative Development and Utilization of Forest Plant Germplasm, Guangzhou 510642, China;
3. Foshan Municipal Forestry Research Institute, Foshan 528222, China;
4. Guangdong Ecology Vocational College, Guangzhou 510520, China
香椿Toona sinensis为楝科Meliaceae香椿属Toona树种,又名椿、椿芽、春甜树、春阳树等。香椿属于落叶乔木,雌雄异株,偶数羽状复叶,圆锥花序,两性花白色,椭圆形蒴果,翅状种子,种子可以繁殖,在我国其分布区北至辽宁,南到海南,西至甘肃,西南至滇黔,东到台湾。香椿为我国特有珍贵速生用材树种,在我国的栽培历史已有2 000多年,山东、河南、安徽、河北等省为香椿主产区。香椿生长迅速,木质坚硬,纹理细致美观,耐水湿,不翘不裂,为家具和建筑的优良用材树种。香椿又是我国著名的药食两用木本植物,其嫩叶营养丰富,可生食、熟食和腌食,叶、根、皮、果实等均可入药。
目前,有关香椿研究报道集中在香椿食用、药用价值和香椿栽培技术等方面[1-3]。有关遗传改良报道较少。孙鸿有等[4]报道了香椿种源试验林的调查结果,有研究者观测了不同种源种子性状的变异[5],开展了不同种源、家系的性状观测与变异分析[6-9]。在香椿苗木生长节律研究方面,仅见林兴春[10]对5个香椿无性系嫁接苗苗期生长节律进行了分析。观测分析香椿不同种源苗期生长节律,对于预测不同种源在栽培地区的适应性和生长潜力,准确掌握苗木生长规律,提高苗木培育质量和加强幼林抚育具有重要意义。本研究从香椿分布区选择有代表性的14个地点采种,对苗木生长进行为期1年的连续观测,并利用Logistic方程拟合其苗高和地径生长过程,划分苗木生长阶段。
1 材料与方法 1.1 材料从我国福建漳州、广东乐昌、广西那坡、广西南宁、广西兴安、贵州独山、湖北建始、湖南常德、湖南凤凰、陕西安康、云南建水、重庆彭水、四川蓬安、甘肃武都和福建漳州共11个省(市、区)14个县采集香椿种子。采种点地理位置和气候条件见表 1。
|
|
表 1 香椿采种点地理位置和气候因子 Table 1 Geographic locations and climatic information of the seed collection sites of Toona sinensis of different provenances |
试验地设在华南农业大学林学院教学科研基地苗圃。苗圃位于北纬23°09′50″,东经113°21′60″,地处南亚热带,属南亚热带典型的季风海洋气候,全年平均气温20~22 ℃,日均气温0 ℃以上,年降水量1 689.3~1 876.5 mm,雨季为4—9月,降水量占全年的85%左右。
1.3 试验设计用于生长节律观测的苗木在2013年秋定植在苗圃。采用完全随机区组设计种植,5个区组,每个区组每个种源种植3株,株行距1 m×1 m。从2014年1月9日—11月23日,每隔15 d(每月的9和24日)测定苗高、地径,共计观测22次。
1.4 生长曲线拟合利用SPSS19.0软件对各种源香椿苗高和地径的生长量作Logistic曲线年生长动态拟合:

|
(1) |
式中,y为苗高或地径的生长量,x为时间,a、b为待定系数,通过SPSS非线性回归分析解出。非线性回归过程初始值赋值时,k设定为任意选择距离最大观测值不远的渐近线的值,a、b用开始2个观测值代入Logistic曲线方程求解得到[11]。
2 结果与分析 2.1 苗高生长节律由图 1可以看出,香椿各种源幼苗高在1—4月为缓慢生长期,5月初—9月底为快速生长期,之后生长趋于缓慢,表现出“慢-快-慢”的生长过程。生长曲线为典型的“S”型曲线。4月中旬之前,各种源间苗高生长差异不大,而进入快速生长期,各种源间苗高生长差异逐渐拉大,至11月,生长最快的四川蓬安种源(pa)与生长最慢的甘肃武都种源(wd)苗高相差162.65 cm,前者为后者的8.06倍。

|
图 1 不同种源香椿苗高累积生长曲线 Figure 1 The growth accumulation curve of Toona sinensis seedling height of different provenance |
从14个种源苗高生长动态曲线(图 2)可以看出,香椿种源苗高生长量集中在6—7月,占测量期生长总量的49.95%。但是,不同种源苗高快速生长的持续时间不同。四川蓬安(pa)、广西那坡(np)、广东乐昌(lcs)等南方种源的快速生长期为4—9月,持续时间长,在8—9月还能快速生长,生长量分别占总量的23.68%、34.87%、18.80%,“S”型生长曲线特别明显。而甘肃武都(wd)、陕西安康(ak)等北方种源的快速生长期集中在6—7月,持续时间短,“S”型生长曲线不明显,整年的苗高生长维持在一个比较低的水平。由此表明,甘肃武都(wd)、陕西安康(ak)等北方种源不适合在广东栽种。

|
图 2 不同种源香椿苗高生长动态 Figure 2 The growth dynamic curve of Toona sinensis seedling height of different provenance |
香椿地径年生长也呈现“慢-快-慢”的生长过程(图 3),香椿各种源幼苗地径1—4月为缓慢生长期,快速生长期从5月初开始,与苗高同期。但快速生长延续至10月中、下旬,之后进入生长缓慢期。持续时间明显长于苗高,整个速生期大约持续5个月左右。5月初之前,不同种源间地径生长差异不大,而从6月初到10月中、下旬,各种源间地径生长差异逐渐拉大,至测量期末,生长最快的四川蓬安种源(pa)与生长最慢的陕西安康种源(ak)地径相差30.96 mm,前者为后者的5.53倍。

|
图 3 不同种源香椿地径累积生长曲线 Figure 3 The ground diameter growth accumulation curve of Toona sinensis seedling of different provenance |
从香椿地径生长动态曲线(图 4)可以看出,香椿地径生长量主要集中在6—8月,占测量期生长总量的53.52%。但不同种源地径快速生长持续的时间不同。四川蓬安(pa)、广西那坡(np)、广东乐昌(lcs)等南方种源在8—9月还能快速生长,生长量分别占测量期生长总量的31.29%、32.85%、30.55%,而北方种源在7月或8月进入缓慢生长期,9月中旬后基本停止生长。

|
图 4 不同种源香椿地径生长动态 Figure 4 The ground diameter growth dynamic curve of Toona sinensis seedling of different provenance |
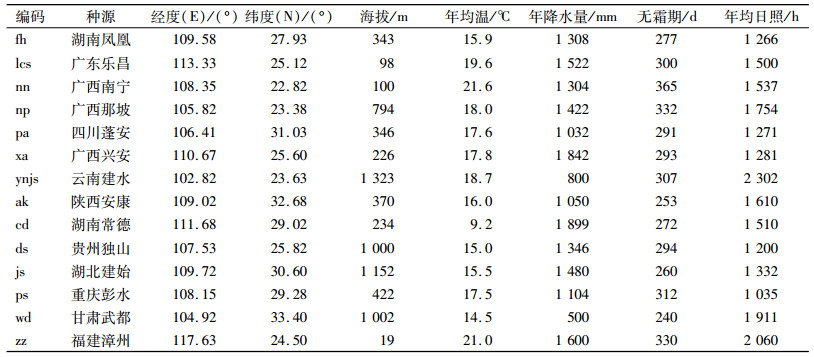
利用Logistic模型拟合香椿各种源的苗高、地径数据,计算出生长曲线模型各参数(表 2)。各种源苗高、地径拟合方程的决定系数(R2)分别为0.945~0.996、0.964~0.994,达到了极显著相关水平。由此说明,用Logistic方程来拟合苗高、地径的生长节律精确度是很高的。
|
|
表 2 不同种源香椿苗高、地径的Logistic回归曲线参数 Table 2 The seedling heights and ground diameters logistic regression parameters of Toona sinensis of different provenances |
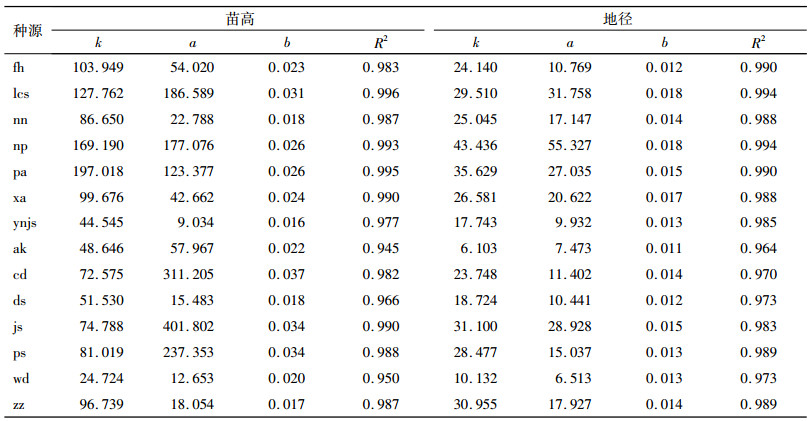
根据不同种源苗高生长曲线的速生拐点、萌动到速生和速生转缓慢的分界点,将不同种源苗高生长划分为3个阶段(表 3)。不同种源苗高生长3个阶段的节律有所不同。苗高生长前期,生长量最大的是四川蓬安种源(pa),为29.97 cm,而生长量最小的云南建水种源(ynjs)为0.45 cm。各种源苗高速生期起始时间通常在4—5月,贵州独山种源(ds)、甘肃武都种源(wd)和云南建水种源(ynjs)速生期起始时间最早,在3月份,主要原因是其整年的生长比较缓慢,以至于生长较缓慢的部分时间划入了速生期,使速生期的起始时间提前1个多月。速生期最晚的是广西那坡种源(np),起始时间在6月8日,主要原因是其整个生长期都生长快速,以至于生长较快的部分时间未划入速生期,使速生期的起始时间较一般情况晚半个月。各种源苗高速生期持续71.2~164.6 d,速生期的苗高生长量为15.62~112.81 cm,速生期占整个测量周期生长量的58.84%~87.83%。后期生长量一般比前期生长量大。
|
|
表 3 各种源香椿苗木不同生长阶段的苗高生长 Table 3 The seedling height growth in different growth stages of Toona sinensis of different provenances |
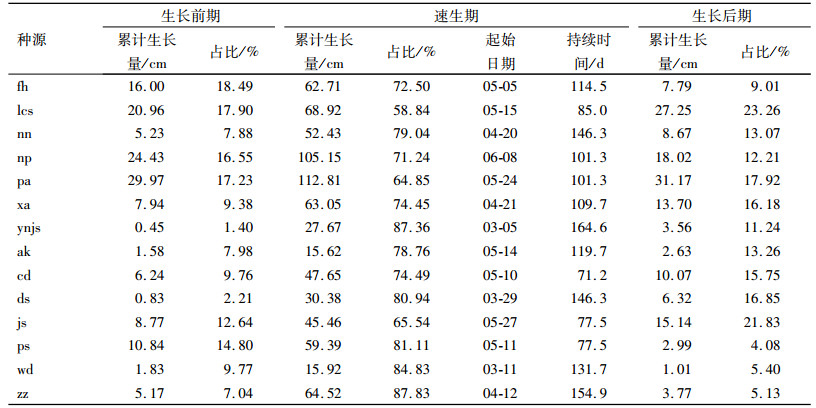
14个香椿种源幼苗地径的生长节律也划分为3个阶段(表 4),地径生长前期,生长量最大的是四川蓬安种源(pa), 生长量为3.92 mm,而生长量最小的陕西安康种源(ak)生长量为0.03 mm。各种源地径速生期起始时间通常在4—5月,与苗高速生期起始时间相当。甘肃武都种源(wd)起始时间最早,在2月,紧随其后的陕西安康种源(ak)、湖南常德种源(cd)、云南建水种源(js),起始时间在3月,最晚的是广西那坡种源(np),速生期起始时间在6月8日。速生期的持续时间为146.3~239.4 d,速生期的地径生长量为4.30~20.87 mm,速生期占整个测量周期生长量的75.94%~97.16%。生长后期生长量一般比生长前期小。
|
|
表 4 各种源香椿苗木不同生长阶段的地径生长 Table 4 The seedling ground diameter growth in different growth stages of Toona sinensis of different provenances |
与苗高生长相比较,同一种源地径速生期起始时间普遍比苗高早。例如,陕西安康种源(ak)和湖南常德种源(cd)的苗高速生期起始时间分别为5月14日和5月10日,而地径速生期起始时间分别为3月13日和3月30日。地径速生期持续时间普遍比苗高长。各种源苗高速生期持续时间平均为114.4 d,而地径速生期持续时间平均为189.2 d。
3 讨论与结论香椿各种源苗高和地径生长节律均呈“S”型,即表现出“慢-快-慢”的生长过程。幼苗高生长在1—4月为缓慢生长期,5月中旬至9月末为快速生长期,之后生长趋于缓慢。而地径生长持续至10月中、下旬。据林兴春[10]对5个香椿无性系8个月嫁接苗苗高和地径生长的调查,苗高和地径的年生长趋势也均为典型的“S”型曲线,苗高和地径的生长主要集中在8月份以前,8月份以后苗高和地径的生长变化不明显。梁有旺等[12]的研究表明,7—9月是香椿苗高生长的主要时期。各地试验结果有关速生期时间有所不同,这可能与地区的气候条件差异有关。
不同种源间苗木生长差异明显。生长最快的四川蓬安种源(pa)与生长最慢的甘肃武都种源(wd)苗高相差162.65 cm,前者为后者的8.06倍。苗高生长主要集中在6—7月,占测量期生长总量的49.95%,南方种源在7月以后的8—9月还能快速生长,四川蓬安种源(pa)、广西那坡种源(np)、广东乐昌种源(lcs)在8—9月的生长量分别占测量期生长总量的23.68%、34.87%、18.80%,而北方的种源则生长缓慢。地径生长最快的四川蓬安种源(pa)与生长最慢的陕西安康种源(ak)地径相差30.96 mm,前者为后者的5.53倍。地径的生长主要集中在6—8月,占测量期生长总量的53.52%,南方种源在9月还能快速生长。从各种源苗木生长节律和总的生长量可以看出,陕西安康(ak)和甘肃武都(wd)等北方种源不适合在广东栽培。
各种源苗高、地径Logistic拟合方程的决定系数分别为0.945~0.996、0.964~0.994,达到了极显著相关水平。多数树种苗木生长节律观测分析结果也表明,用Logistic方程来拟合苗高、地径的生长节律有很高的精确度[13-15]。本研究的各种源苗高速生期起始时间集中在4—5月。速生期的持续时间为71.2~164.6 d,速生期的生长量占整个测量期的58.84%~87.83%。生长后期生长量一般比生长前期大。各种源地径速生期起始时间集中在4—5月,与苗高速生期起始时间相当,持续时间为146.3~239.4 d。
林兴春[10]观测结果表明,香椿嫁接后地径比苗高更早进入速生期,地径的速生期也比苗高长。这与本研究结论是一致的。通过对香椿苗木年生长节律的观测研究,对其生长期进行划分,可针对不同的生长阶段,制定不同的栽培技术措施。速生期是植株生长的关键期,速生期的长短决定了其当年生长量[16],应抓住速生期,加强水肥管理,促进香椿的快速生长。
| [1] |
SU Y F, YANG Y C, HSU H K, et al. Toona sinensis leaf extract has antinociceptive effect comparable with non-steroidal anti-inflammatory agents in mouse writhing test[J]. BMC Comp Altern Med, 2015, 15(1): 1-4. DOI:10.1186/s12906-015-0520-z (  0) 0) |
| [2] |
XIA Q, WU W C, TIAN K, et al. Effects of different cutting traits on bud emergence and early growth of the Chinese vegetable Toona sinensis[J]. Sci Hortic, 2015, 190: 137-143. DOI:10.1016/j.scienta.2015.04.026 ( 0) |
| [3] |
ZHANG B B, MA Z Q, ZHANG J, et al. Effects of rehydration conditions on quality of dehydrated Toona sinensis[J]. Mod Food Sci Technol, 2015, 31(7): 270-276. ( 0) |
| [4] |
孙鸿有, 王鹏飞, 方炳法, 等. 香椿地理变异与种源选择[J]. 浙江林学院学报, 1992, 9(3): 4-12. ( 0) |
| [5] |
周祥斌, 周玮, 周鹏, 等. 香椿种源间种子性状地理变异研究[J]. 华南农业大学学报, 2015, 36(5): 105-110. ( 0) |
| [6] |
李淑玲, 桑玉强, 王平, 等. 不同种源香椿性状遗传分析[J]. 河南农业大学学报, 2000, 34(4): 363-366. DOI:10.3969/j.issn.1000-2340.2000.04.015 ( 0) |
| [7] |
潘晓芳, 宋志姣, 陶思叁, 等. 广西香椿种源种苗特性研究[J]. 广东农业科学, 2013, 40(4): 20-22. DOI:10.3969/j.issn.1004-874X.2013.04.007 ( 0) |
| [8] |
郝明灼, 陈德根, 彭方仁, 等. 8个种源香椿种子性状及芽苗菜产量和品质比较[J]. 浙江农林大学学报, 2012, 29(2): 80-184. ( 0) |
| [9] |
刘军, 陈益泰, 姜景民, 等. 香椿种源苗期性状变异与原产地生态因子典型相关分析[J]. 东北林业大学学报, 2010, 38(11): 27-29. DOI:10.3969/j.issn.1000-5382.2010.11.008 ( 0) |
| [10] |
林兴春. 5个香椿无性系嫁接苗苗期生长节律分析[J]. 林业勘察设计, 2015(1): 124-127. DOI:10.3969/j.issn.1004-2180.2015.01.030 ( 0) |
| [11] |
董江水. 应用SPSS软件拟合Logistic曲线研究[J]. 金陵科技学院学报, 2007, 23(1): 21-24. ( 0) |
| [12] |
梁有旺, 彭方仁, 陈德平. 不同种源香椿苗期生长差异比较[J]. 林业科技开发, 2007, 21(2): 38-41. DOI:10.3969/j.issn.1000-8101.2007.02.012 ( 0) |
| [13] |
邝雷, 邓小梅, 陈思, 等. 4个任豆种源苗期生长节律的研究[J]. 华南农业大学学报, 2014, 35(5): 98-101. ( 0) |
| [14] |
陈丽君, 刘明骞, 廖柏勇, 等. 苦楝不同种源苗期生长性状和生长节律研究[J]. 西南林业大学学报, 2014, 34(4): 1-7. DOI:10.3969/j.issn.2095-1914.2014.04.001 ( 0) |
| [15] |
廖海红, 孔小丽, 周华, 等. 毛红椿九连山种源播种苗苗期生长规律研究[J]. 南方林业科学, 2015, 43(1): 11-16. ( 0) |
| [16] |
彭玉华, 黄志玲, 申文辉, 等. 台湾桤木引种的生长节律与生物量研究[J]. 2014, 34(6): 7-12. http://kns.cnki.net/KCMS/detail/detail.aspx?filename=znlb201406003&dbname=CJFD&dbcode=CJFQ
( 0) |